(Primera editorial: https://wp.me/pbW3AH-1H7)
Traducido al español por Carmen Sanabria Ayala
Entre las consecuencias está la inhibición del transporte de vitamina B9 (folato) al sistema nervioso, con profundas implicaciones terapéuticas que, a pesar de estar documentadas, han sido en gran medida ignoradas. Segundo Editorial de este Site sobre las causas del autismo.
Fundamentándose en la lógica iluminada por las publicaciones científicas que se han ido acumulando a lo largo de décadas, esta secuencia de Editoriales pretende ofrecer al lector un camino hacia la identificación de la etiología (causa) del autismo, una enfermedad cuyo coste anual entre gastos directos e indirectos puede alcanzar más de 460 mil millones de dólares en EE.UU. en 2025 (https://pubmed.ncbi.nlm.nih.gov/26183723/).
El editorial anterior reunió datos clínicos y científicos que identifican la naturaleza inflamatoria del proceso patológico que causa el comportamiento autista. Se excluye así de forma incuestionable (a través de un simple examen de laboratorio, como es la medición de los niveles circulantes de la enzima enolasa neuronal específica) la hipótesis de que el niño autista sería simplemente un individuo “neuroatípico”, – término que erróneamente lo considera simplemente “diferente” y no como portador de déficits de carácter orgánico, o sea, portador de una enfermedad física). En este sentido, es necesario recordar que hasta el 60% de los autistas presentan deficiencia cognitiva (https://www.sciencedirect.com/science/article/abs/pii/S0006322396005227).
Así, la caracterización de una encefalitis crónica, con liberación de la mencionada enzima en la circulación por las células neuronales lesionadas, es suficientemente elocuente para la caracterización de una enfermedad neurológica. La persistencia en negar este hecho sólo perjudica la identificación y aplicación oportuna de un tratamiento temprano y eficaz – el derecho del propio niño y de sus padres. Al reconocerse el autismo como un trastorno neurológico, la identificación de su causa fundamental conduciría al desarrollo de estrategias preventivas, deteniendo la actual epidemia de autismo, lo que sería el mejor interés para la sociedad en su conjunto (incluida la comunidad médica). Como se verá en el próximo Editorial, la epidemia de autismo ya no puede considerarse meramente aparente o resultado de “mejores diagnósticos y concientización” (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6242891/) lo que llevó a acuñar la expresión “Trastornos del Espectro Autista” (TEA) en lugar de “autismo”. De hecho, ¿cómo es posible creer que un simple trastorno de conducta preexistente, cuya prevalencia no estaría aumentando (sólo sería mejor “percibida” o “detectada”), tenga una previsión de generar un costo anual de más de 460 billones de dólares en EE.UU. para el 2025 (https://pubmed.ncbi.nlm.nih.gov/26183723/)?
Un proceso inflamatorio que afecta al cerebro (encefalitis) puede ser causado, por ejemplo, por infecciones virales o bacterianas o incluso por una agresión del sistema inmunológico (la encefalitis autoinmune). Además, hay un hecho muy importante en lo que se refiere al autismo (que será revisado en posteriores Editoriales de este site): la encefalitis crónica que caracteriza esa condición también puede ser causada por la presencia de aluminio en el tejido nervioso (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10180736/) desencadenando un proceso autoinmune dirigido contra proteínas (antígenos) neuronales.
Este Segundo Editorial se centra en la agresión autoinmune crónica presente en los portadores de autismo (encefalitis crónica). La avalancha de datos científicos demostrativos de la presencia de un proceso autoinmune en esa condición contrasta con la visión de que se trata de un simple trastorno conductual (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5373490/). Como veremos en editoriales posteriores, en consonancia con el consagrado “Principio de Parsimonia”, inspirador de la estadística moderna, es necesario identificar el factor desencadenante primordial de esa agresión autoinmune, alrededor del cual gravitan diversos factores secundarios que contribuyen para su progresión y sustentación. Este principio se utiliza, por ejemplo, en la práctica médica diaria para establecer un único diagnóstico (el más probable), en contraposición a la improbabilidad de que se produzcan múltiples diagnósticos simultáneamente en un solo paciente. En la identificación del diagnóstico de la enfermedad, es decir, de la causa única de un conjunto de manifestaciones clínicas y de laboratorio, siempre se debe buscar la explicación más simple (siempre que sea posible, la única) para todas las manifestaciones.
Así, de la misma forma como es extremadamente improbable que un paciente presente dos enfermedades que se inician simultáneamente sin ninguna relación fisiopatológica entre sí, es también altamente improbable que las piezas del rompecabezas fisiopatológico del autismo no se encajen perfectamente en torno de un factor causal primordial.
De la misma forma todos los descubrimientos que han sido relatados en el autismo, incluyendo su condición de encefalitis crónica, las manifestaciones autoinmunes, los factores genéticos predisponentes y, especialmente, el crecimiento exponencial de su incidencia y prevalencia que se ha producido en los últimos 40-50 años, tienen que ser agruparse y ordenarse en torno a una de esa causa principal, como montar las piezas de un rompecabezas. Nuevamente se enfatiza aquí que, sin identificar esta causa primaria, no hay manera de viabilizar la interrupción del proceso patológico (inflamatorio, autoinmune, neurodegenerativo) que caracteriza al autismo o implementar su prevención.
La afirmación de que “el autismo tiene múltiples causas que ocurren en diversas combinaciones” o que se trata de “un disturbio biológico altamente complejo y heterogéneo” (https://www.biologicalpsychiatryjournal.com/article/S0006-3223(16)32739-1/) va en contra del Principio de la Parsimonia (o “navaja de Occam”). Las afirmaciones sólo sirven para desvanecer la búsqueda de la causa primordial de esta tragedia humanitaria, social y económica, mientras que su prevalencia sigue en rápido ascenso en ausencia de medidas preventivas que se oponen a ella. Por el contrario, en esta secuencia de editoriales buscamos equiparar la diversidad de hallazgos fisiopatológicos relacionados con el autismo con las diversas manifestaciones clínicas y de laboratorio encontradas en un paciente, de forma a identificar una única causa primordial –comparativamente, un diagnóstico único frente a múltiples diagnósticos.
Por otro lado, la afirmación de que “una persona con autismo nace con autismo” (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5501015/), en particular, desconsidera el hecho actualmente indiscutible de que muchos niños desarrollan las primeras manifestaciones del autismo después del primer año de vida, después de haber tenido un desarrollo psicomotor inicial perfectamente normal (“autismo regresivo”, – que fue negado de forma especulativa por demasiado tiempo). (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4949854/).
Sin embargo, el autismo regresivo realmente existe (https://goldencaretherapy.com/regressive-autism – 2021):
“El autismo regresivo, también conocido como autismo de aparición tardía,
implica un período de desarrollo típico seguido por una pérdida de
habilidades previamente adquiridas o una disminución perceptible de las habilidades sociales y de comunicación. Esta regresión generalmente ocurre entre los 15 y 30 meses de edad y puede ser repentina o gradual”.
El siguiente ejemplo fue tomado de un texto escrito hace algunos años. El autor intentaba en esa época (2017) argumentar contra la realidad de la existencia de casos de autismo regresivo (https://www.thetransmitter.org/spectrum/rethinking-regression-autism/):
“…un niño de 2 años, hablador y curioso, de repente se retrae. Se vuelve indiferente al sonido de su nombre. Él empieza a hablar menos que antes o deja de hablar completamente. Deja de jugar con las personas y pasa a jugar con cosas, de explorar muchos objetos y actividades hasta obsesionarse por algunos. Pierde muchas de las habilidades que dominaba y comienza a balancearse, girar, andar de puntillas o dar palmadas. Es frecuentemente en este punto, que sus padres aterrorizados buscan respuestas de especialistas”.
… y los padres normalmente no encuentran respuestas asertivas coherentes con la lógica científica, sino sólo especulaciones. Ese hecho es debido a la falta de una presentación integrada y racionalizada de las características fisiopatológicas ya documentadas a través de diferentes investigaciones. Esta brecha es precisamente el objetivo que inspiró estos editoriales consecutivos que también deben explicar la existencia del autismo regresivo.
Contrariamente a los intentos de negar su existencia, la realidad del autismo regresivo se encuentra desde hace muchos años reconocida en publicaciones científicas, donde los autores lo señalan como “intrigante” (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4949854/), reconociendo no saber cómo explicarlo:
“La aparición de regresión del desarrollo en el trastorno del espectro autista (TEA) es uno de los fenómenos más intrigantes de ese trastorno”.
“To date, the causes of regression in autism are unknown.”
En este Editorial revisamos el impacto directo de esta agresión autoinmune sobre el acceso de un nutriente de vital importancia al Sistema Nervioso Central (SNC): la vitamina B9 (o “folato”: metilfolato y ácido folínico – este último también llamado de leucovorina). La presencia de polimorfismos genéticos (genes con una estructura diferente a la conformación padrón) tornan una fracción muy significativa de la población infantil particularmente sensible a la reducción del aporte de los folatos al SNC.
La agresión autoinmune al cerebro del niño autista se confirma a través de reiteradas publicaciones iniciadas en la década de 1980 y posteriormente revisadas por varios autores (https://www.sciencedirect.com/science/article/pii/S1750946720300581; https://www.frontiersin.org/journals/cellular-neuroscience/articles/10.3389/fncel.2018.00405/full#B90; https://psychiatryonline.org/doi/10.1176/appi.focus.24022004; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8955336/; https://www.nature.com/articles/npp2016158; https://psychiatryonline.org/doi/10.1176/appi.focus.24022004;
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6242891/; https://pubmed.ncbi.nlm.nih.gov/15223250/;
Las evidencias de que la encefalitis crónica que afecta al niño autista implica una agresión autoinmune al tejido cerebral puede ser organizada de la siguiente manera:
(I) Generación de autoanticuerpos dirigidos contra antígenos neurales;
(II) Generación de autoanticuerpos dirigidos contra antígenos no específicamente neurales
(III) Correlación entre los niveles de autoanticuerpos circulantes y la gravedad del autismo;
(IV) Presencia de linfocitos infiltrando el cerebro de individuos con autismo;
(V) La respuesta inmune Th17 (inducida por linfocitos auxiliares productores de interleucina 17, típica en condiciones autoinmunes) fue observada en individuos autistas;
(VI) Atenuación de los síntomas autistas mediante terapia con corticosteroides o terapia con inmunoglobulina;
(VII) Aparición de trastornos autoinmunes o inmunomediados como comorbilidades en el autismo;
(VIII) Prevalencia elevada de enfermedades autoinmunes entre parientes consanguíneos (un historial familiar de trastornos autoinmunes aumenta el riesgo de autismo);
(IX) Al igual que en las enfermedades autoinmunes clásicas, los beneficios de la administración de colecalciferol también son observados en el autismo;
(X) Demostración de una resistencia genética a los efectos inmunorreguladores del colecalciferol (o “vitamina” D, un inhibidor de la actividad Th17) en el autismo.
Sigue a continuación la presentación de cada grupo de publicaciones.
(I) Generación de autoanticuerpos dirigidos contra antígenos neurales.
El sistema inmune de los portadores de autismo genera una gran variedad de autoanticuerpos dirigidos contra antígenos neurales (https://www.frontiersin.org/journals/cellular-neuroscience/articles/10.3389/fncel.2018.00405/full#B90; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6952169/), con reactividad dirigida contra diversas regiones del SNC y diferentes poblaciones de neuronas. En conjunto con los otros datos publicados que se enumeran más abajo, la producción de autoanticuerpos no puede ser considerada simplemente un epifenómeno del proceso que causa el autismo; por el contrario, debe ser considerada un factor cardinal en el proceso inflamatorio nocivo para el SNC (un epifenómeno sería un fenómeno asociado a un proceso lesional sin una relación causal con él).
De acuerdo con el Principio de la Parsimonia, la causa primaria del autismo debe explicar tanto el amplio repertorio de autoanticuerpos producidos como la variedad de regiones y células neurales afectadas. Como se mencionó en la introducción a este Editorial, la causa primordial, en torno a la cual gravitan factores agravantes, será resaltada en un próximo editorial, aplicando los criterios de causalidad establecidos por Austin Bradford Hill en 1965 y consagrados desde entonces.
La presencia de autoanticuerpos dirigidos contra proteínas del SNC en individuos autistas fue identificada por primera vez por Singh y colaboradores en 1988. Ellos encontraron anticuerpos contra proteínas de filamentos de axones neuronales (NAFP) en la sangre de 10 de 15 niños portadores de autismo (https://pubmed.ncbi.nlm.nih.gov/3144935/). Los anticuerpos antiproteína básica de mielina (MBP) fueron identificados en individuos con TEA en 1993 (https://pubmed.ncbi.nlm.nih.gov/7682457/) – hallazgo que fue replicado en 1998 (https://pubmed.ncbi.nlm.nih.gov/9756729/) y en el 2006 (https://pubmed.ncbi.nlm.nih.gov/16181614/). La presencia de anticuerpos anti-NAFP también fue confirmada en 1998 (https://pubmed.ncbi.nlm.nih.gov/9756729/). En el 2013 niveles significativamente elevados de anticuerpos anti-MBP fue confirmada en comparación con controles sanos (https://pubmed.ncbi.nlm.nih.gov/23726766/); en el mismo estudio, niveles significativamente elevados de anticuerpos anti-glicoproteína asociada a la mielina (“anti-myelin associated glycoprotein”, “anti-MAG”) fueron demostrados en comparación con controles sanos. El mismo estudio demostró niveles significativamente elevados de anticuerpos anti-glicoproteína asociada a la mielina (“anti-myelin associated glycoprotein”, “anti-MAG”) en comparación con controles sanos. El mismo estudio demostró también la vinculación de estos autoanticuerpos a la gravedad del autismo:
“Los pacientes con autismo grave presentaron autoanticuerpos séricos anti-MBP y anti-MAG significativamente más elevados que los niños con autismo leve a moderado, P = 0,047 y P <0,001, respectivamente (Tablas 1 y 2)”.
Otros estudios demostraron un aumento significativo en la incidencia de anti-NAFP y anti-proteína ácida fibrilar glial (GFAP) en individuos autistas, pero no en individuos con retraso mental (https://www.sciencedirect.com/science/article/abs/pii/S0887899497000453).
Estudios más recientes han encontrado autoanticuerpos dirigidos a las regiones de la corteza prefrontal, caudado, putamen, cerebelo y giro cingular del cerebro (https://pubmed.ncbi.nlm.nih.gov/16842863/) e hipotálamo (https://pubmed.ncbi.nlm.nih.gov/17804536/) en niños con TEA.
Según las conclusiones de los autores de este último estudio:
“Aunque el papel potencial de estos autoanticuerpos en el autismo, actualmente sea desconocido, su presencia sugiere una pérdida de autotolerancia a uno o más antígenos neuronales durante la primera infancia”.
De la misma forma, en otro estudio publicado en 2009 (https://pubmed.ncbi.nlm.nih.gov/18706993/), Los investigadores encontraron que el 21% de las muestras de plasma de niños con TEA eran altamente inmunorreactivas contra las neuronas de Golgi de los primates. Estas interneuronas inhibidoras están localizadas en la capa granular del cerebelo. Ellos utilizan el neurotransmisor ácido gamma-aminobutírico (GABA) para modular las sinapsis excitadoras, permitiendo un equilibrio entre excitación e inhibición. Por el contrario, esta inmunorreactividad no se observó cuando fue usado el plasma obtenido de controles con desarrollo típico y de la misma edad.
Un estudio posterior identificó que la autorreactividad tenía también como objetivo otras interneuronas GABAérgicas distribuidas por toda el neocórtex y muchas regiones subcorticales, incluyendo las capas superficiales del córtex (https://pubmed.ncbi.nlm.nih.gov/21521495/). Otros autores verificaron que la inmunorreactividad dirigida a las neuronas de Golgi y otras interneuronas se correlaciona con la gravedad de las alteraciones conductuales y emocionales en los niños autistas (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3313674/; https://pubmed.ncbi.nlm.nih.gov/21420487/;https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3039058/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4111628/; https://pubmed.ncbi.nlm.nih.gov/22226851). Según con el resultado de estos estudios, las alteraciones en la estructura del cerebelo y en la composición de la población de neuronas cerebelosas, se encuentran entre las anomalías más consistentes del autismo (https://pubmed.ncbi.nlm.nih.gov/11468308/).
Al contrario de lo que se pensaba, las células del sistema inmunológico penetran rutinariamente el SNC incluso en ausencia de inflamación con el propósito de vigilancia inmunológica y la presentación local de antígenos con la finalidad de producción de anticuerpos y proliferación de clones linfocitarios, de hecho ocurre incluso en sistema nervioso no inflamado (https://www.nature.com/articles/nn.3161).
En ocasiones, de la neuroinflamación se rompen las barreras que separan el encéfalo de la circulación, aumentando el tránsito de células y macromoléculas entre estos compartimentos (https://www.mdpi.com/1422-0067/24/16/12699). Por lo tanto, la penetración de autoanticuerpos dirigidos contra las neuronas GABAérgicas puede reducir el número o la actividad de esas células inhibitorias. Este hecho puede contribuir para el desequilibrio entre excitación e inhibición, – un desequilibrio que durante mucho tiempo se ha identificado como determinante de la perturbación de los sistemas sensoriales, mnemotécnicos, sociales y emocionales que se encuentran en el autismo https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6748642/).
Se conoce desde hace mucho tiempo que las células gliales (que ayudan a aislar, sostener y nutrir las neuronas) se multiplican (mediante un proceso llamado mitosis).
Al contrario de conceptos más antiguos, la generación de nuevas neuronas también fue demostrada en el SNC humano adulto, desde hace más de un cuarto de siglo, aunque no mediante mitosis. Las neuronas recién formadas surgen de la multiplicación de células localizadas alrededor de las cavidades y canales a través de los cuales circula el líquido cefalorraquídeo — la capa “subependimaria”. El término “epéndimo” se utiliza para indicar la capa de células que recubren las cavidades internas del cerebro (ventrículos), mientras que la región “subependimaria” denota el área adyacente a los ventrículos. Las células de esta región se pueden clasificar en dos tipos: células madre neurales y células progenitoras neurales, estas últimas derivadas de células madre neurales. Cuando se multiplican, ambas dan lugar a la formación de nuevas neuronas (neurogénesis) y dos tipos de células gliales: astrocitos y oligodendrocitos, que constituyen la “macroglia”. Ellas participan de la formación y maduración del sistema neuronal, no sólo en las etapas embrionaria y fetal, sino también después del nacimiento, y la producción de nuevas neuronas continúa incluso en individuos de edad avanzada. Esas neuronas recién formadas (“neuroblastos”) son capaces de migrar a partir de la región subependimaria para varias regiones del SNC, apoyando la población de células neurales y la función de esas regiones (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6782600/; https://www.jneurosci.org/content/22/3/612.short; https://www.nature.com/articles/nm1198_1313; https://www.sciencedirect.com/science/article/pii/S2214854X20300133). La neurogénesis probablemente esté involucrada en los procesos de adquisición de habilidades en la infancia, como el habla (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3844860/). Por otro lado, el desarrollo normal del sistema nervioso en el periodo posnatal requiere que sea eliminado un exceso de conexiones (sinapsis) entre neuronas, mediante un proceso denominado “poda sináptica” (https://www.science.org/doi/abs/10.1126/science.1202529). Ese proceso se encuentra inhibido en el cerebro del portador de autismo (https://www.nature.com/articles/mp2016103).
Un alto nivel sérico de autoanticuerpos generados contra células progenitoras neurales humanas fue identificado en pacientes con autismo (https://pubmed.ncbi.nlm.nih.gov/23838310/). Por otro lado, el suero de pacientes con autismo suprime la diferenciación y la maduración de células progenitoras neuronales en cultivo, demostrando un evento autoinmune que puede ser uno de los mecanismos de deterioro del neurodesarrollo en esa condición, involucrando la inhibición de la neurogénesis (https://pubmed.ncbi.nlm.nih.gov/19526302/; https://pubmed.ncbi.nlm.nih.gov/19526302/; https://pubmed.ncbi.nlm.nih.gov/23838310/).
Por otro lado, la excesiva activación de la microglía, como parte del mecanismo implicado en la inflamación que afecta al cerebro autista (https://www.sciencedirect.com/science/article/abs/pii/S0022395620308785), puede alterar la ejecución del proceso de poda neural por parte de esas células, lo que lleva a un exceso de sinapsis excitadoras en detrimento de las sinapsis inhibidoras (https://www.sciencedirect.com/science/article/abs/pii/S0168010215001625), contribuyendo así, a través también de ese mecanismo, para una hiperexcitación potencialmente subyacente a la hiperactividad encontrada en el autista.
Contribuyendo para ese desequilibrio, se verifica la producción de autoanticuerpos dirigidos contra los receptores de la serotonina en el cerebro autista (https://pubmed.ncbi.nlm.nih.gov/2578670/; https://pubmed.ncbi.nlm.nih.gov/1375597/). La serotonina es un neurotransmisor de fundamental importancia para el comportamiento emocional y el deterioro de la actividad serotoninérgica puede alterar la atención y las reacciones emocionales ante estímulos externos (https://www.sciencedirect.com/science/article/abs/pii/S1569733910700904), tal como se verifica en el comportamiento autista (https://www.sciencedirect.com/science/article/abs/pii/S0890856713003080). Los niveles séricos de anticuerpos dirigidos contra el receptor de serotonina en un niño autista alcanzaron valores de 600 y 980 fmol/ml de suero en dos muestras recogidas con un mes de diferencia. Estos niveles son muy superiores a los niveles de anticuerpos dirigidos contra los receptores nicotínicos del neurotransmisor acetilcolina, presente en una enfermedad clásicamente reconocida como autoinmune (la miastenia gravis), en la que los títulos de autoanticuerpos séricos alcanzan valores máximos de 45 fmol/ml, determinando reducción de la fuerza del músculo esquelético (https://pubmed.ncbi.nlm.nih.gov/2578670/).
Entre los autoanticuerpos dirigidos contra las células cerebrales encontrados en el suero de los niños autistas , se encuentran también los anticuerpos anti-glicoproteína básica asociada a la mielina (anti-MAG) (https://pubmed.ncbi.nlm.nih.gov/22898564/;https://pubmed.ncbi.nlm.nih.gov/23726766/), anti-proteína mielínica básica (anti-MBP) (https://pubmed.ncbi.nlm.nih.gov/23726766/) y anticuerpos anti-gangliósido M1 (el componente glicoesfingolípido más abundante de las membranas neuronales). Los niveles más elevados de estos autoanticuerpos son encontrados en los portadores de cuadros más severos de autismo (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3104945/).
Otros autoanticuerpos cerebrales específicos, como anticuerpos anti-proteína básica de la mielina (https://pubmed.ncbi.nlm.nih.gov/15223250/), anticuerpos anti-glicoproteína asociada a la mielina y anticuerpos anti-gangliósido M1 (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3104945/), se observaron en niños autistas.
(II) Generación de autoanticuerpos dirigidos contra antígenos no específicamente neuronales
El factor antinuclear (ANA), también conocido como ANA (anticuerpo antinuclear), está compuesto por anticuerpos dirigidos contra estructuras (antígenos) localizadas en el núcleo y en el citoplasma de las células, como proteínas, ácidos nucleicos (ADN, ARN) y complejos de proteínas y ácidos nucleicos. Su presencia en la circulación ha sido considerada hace décadas como fundamental para el diagnóstico de enfermedades autoinmunes (https://ard.bmj.com/content/73/1/17.short; https://www.sciencedirect.com/science/article/abs/pii/S2173574310700496). Su producción por las células del sistema inmunológico de un individuo (también en niños) indica que una enfermedad autoinmune puede estar presente o en desarrollo, como lupus eritematoso sistémico (LES), esclerodermia (localizada o sistémica), enfermedad mixta del tejido conjuntivo (con características mixtas de LES y polimiositis), artritis reumatoide, artritis reumatoide juvenil, síndrome de Sjögren, polimiositis y dermatomiositis (https://my.clevelandclinic.org/health/diagnostics/14897-antinuclear-antibody-test-in-children;https://www.researchgate.net/publication/333705452_Pattern_and_Frequency_of_Anti-nuclear_Antibody_Positivity_in_Paediatric_Rheumatic_Diseases; https://www.sciencedirect.com/science/article/pii/S2589909022000065).
Como sería de esperar en el caso de la participación de mecanismos autoinmunes en el autismo, se encontró un resultado positivo en el test de FAN (ANA) en niños autistas asociado con la gravedad de las manifestaciones autistas y la presencia de alteraciones electroencefalográficas (https://pubmed.ncbi.nlm.nih.gov/19135624/).
Los anticuerpos antifosfolípidos son autoanticuerpos que tienen como objetivo proteínas unidas a fosfolípidos (estructuras lipídicas fundamentales de la membrana de las células) (https://pt.khanacademy.org/science/ap-biology/cell-structure-and-function/plasma-membranes/a/structure-of-the-plasma-membrane#:~:text=v%C3%AAm%20dos%20carboidratos.-,Fosfolip%C3%ADdios,t%C3%AAm%20regi%C3%B5es%20hidrof%C3%ADlicas%20e%20hidrof%C3%B3bicas). La presencia de estos anticuerpos conduce al síndrome del anticuerpo antifosfolípido (SAF, una enfermedad autoinmune multisistémica – https://pubmed.ncbi.nlm.nih.gov/36849186/). El resultado es un riesgo aumentado de eventos trombóticos, morbilidad durante el embarazo y muchas otras complicaciones autoinmunes e inflamatorias (https://pubmed.ncbi.nlm.nih.gov/36849186/). A pesar de haber sido descrita inicialmente en el cuadro de lupus (LES), el SAF también se encuentra disociado del LES con frecuencia similar (https://pubmed.ncbi.nlm.nih.gov/36849186/). Además del LES, se encuentran asociadas al SAF enfermedades como la trombocitopenia, anemia hemolítica, la valvulopatía, la hipertensión pulmonar, la nefropatía microangiopática, úlceras cutáneas, livedo reticularis, migraña refractaria, disfunción cognitiva y la aterosclerosis (https://www.ncbi.nlm.nih.gov/books/NBK459442/). Apoyando la implicación de mecanismos autoinmunes en el autismo, niveles elevados de autoanticuerpos antifosfolípidos (anticardiolipina, anti-β2-glicoproteína 1 y antifosfoserina) han sido encontrados en individuos con autismo y están asociados con la gravedad de las alteraciones del comportamiento (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3794552/).
Los anticuerpos anticélulas endoteliales son un grupo heterogéneo de anticuerpos dirigidos contra las células de revestimiento interno de los vasos sanguíneos. Desde su descubrimiento en la década de 1970, estos autoanticuerpos han sido identificados en diferentes condiciones caracterizadas por inflamación vascular, incluyendo LES, SAF, vasculitis sistémica, artritis reumatoide, esclerodermia sistémica y rechazo de órganos trasplantados (https://www.sciencedirect.com/science/article/abs/pii/S1568997216302798). De forma similar a lo que ocurre en relación con el FAN y a los anticuerpos antifosfolípidos, la presencia de niveles más elevados de anticuerpos anti células endoteliales circulantes, tiene una relación directa con la gravedad de las alteraciones del comportamiento en portadores de autismo (https://www.sciencedirect.com/science/article/pii/S1726490115000969?via%3Dihub).
La detección de anticuerpos antimitocondriales (AMA) se utiliza para diagnosticar la enfermedad autoinmune llamada colangitis biliar primaria. También puede ocurrir en otras enfermedades autoinmunes, como el síndrome de Sjögren, la esclerosis sistémica (o esclerodermia), la polimiositis/dermatomiositis, la artritis idiopática juvenil, el LES y la hepatitis autoinmune (https://link.springer.com/article/10.1007/s12016-021-08904-y; https://www.sciencedirect.com/science/article/pii/S2589909022000065; https://www.sciencedirect.com/science/article/abs/pii/S0889857X05702970). Los anticuerpos anti-ADN de doble cadena son considerados marcadores altamente específicos del LES (https://www.nature.com/articles/s41584-020-0480-7) y de la hepatitis autoinmune lupoide (https://www.nature.com/articles/s41584-021-00573-7).
Las presencias de AMA (https://jneuroinflammation.biomedcentral.com/articles/10.1186/1742-2094-7-80; https://pubmed.ncbi.nlm.nih.gov/24837704/)), anticuerpos anti-ADN de doble cadena, anticuerpos antinucleares (https://pubmed.ncbi.nlm.nih.gov/24837704/) y anticuerpos antinucleosomas (https://pubmed.ncbi.nlm.nih.gov/24708718/) también fueron documentados en el suero de individuos autistas (los nucleosomas son unidades estructurales que forman los cromosomas y están compuestos por dos espirales de ADN enrolladas alrededor de un disco de proteína que consiste de cuatro pares de proteínas llamadas histonas). La prueba de anticuerpos antinucleótidos se considera altamente sensible y específico para el diagnóstico de LES (https://pubmed.ncbi.nlm.nih.gov/20374326/). Esto indica que la autoinmunidad encontrada en el autismo, que es parte del proceso inflamatorio descrito en el primer editorial de esta serie, puede no ser solo específica de un órgano en muchos casos.
En otras palabras, puede que no esté dirigido únicamente contra el cerebro o el SNC del individuo autista, sino más sí una condición en la que el sistema inmunológico ataca a otros órganos o sistemas.
Nuevamente, siguiendo el Principio de Parsimonia, según el cual la explicación más simple y completa debe considerarse como la más probable de cualquier fenómeno (incluyendo cualquier enfermedad), la búsqueda de la causa determinante primaria del autismo debe necesariamente identificar como causal un factor que sea capaz de explicar no solamente la agresión contra el SNC, sino también la agresión autoinmune dirigida contra los otros órganos y sistemas.
(III) Correlación entre los niveles de autoanticuerpos circulantes y la gravedad del autismo
Los niveles de autoanticuerpos se han identificados como marcadores de la actividad y gravedad de las enfermedades autoinmunes (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC209428/).
Corroborando la participación del fenómeno autoinmune en la fisiopatología del autismo, esa correlación (relación directa) ha sido documentada en niños autistas
(https://www.sciencedirect.com/science/article/pii/S1726490115000969?via%3Dihub; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3794552/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3039058/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3104945/; https://pubmed.ncbi.nlm.nih.gov/22226851; https://pubmed.ncbi.nlm.nih.gov/19135624/).
(IV) Presencia de linfocitos infiltrando en el cerebro de individuos con autismo;
La acumulación de linfocitos se encuentra presente en los tejidos y órganos afectados por enfermedades autoinmunes. En asociación con la inflamación y con la producción de autoanticuerpos, este hallazgo (encontrado en biopsias o autopsias analizadas al microscopio) es considerado la marca registrada de las enfermedades autoinmunes (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2832720/; https://pathology.jhu.edu/autoimmune/damage).
De la misma forma, infiltrados linfocíticos fueron encontrados en el cerebro de los portadores de autismo (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7210715/).
Junto con las evidencias del proceso inflamatorio encefálico en actividad (encefalitis crónica: primer Editorial de esta serie) y del repertorio de anticuerpos producidos contra el tejido encefálico, aquí enumerado, la presencia de linfocitos autorreactivos infiltrando el tejido nervioso del autista finaliza la caracterización de la tríada clásicamente considerada la marca registrada de la agresión autoinmune (https://www.sciencedirect.com/science/article/abs/pii/S0091674909014602) como partícipe del mecanismo fisiopatológico del autismo.
Por otro lado, el descubrimiento de vasos linfáticos activos conectando el SNC al sistema linfático (https://www.nature.com/articles/npp2016158; https://pubmed.ncbi.nlm.nih.gov/26077718/) indica un posible canal de comunicación a través del cual la agresión autoinmune puede actuar en el cerebro autista y otras enfermedades neurológicas autoinmunes.
(V) La respuesta inmune Th17 (inducida por linfocitos auxiliares productores de interleucina 17, típica en condiciones autoinmunes –https://link.springer.com/article/10.1007/s00281-019-00733-8) fue observada en individuos autistas (https://link.springer.com/article/10.1186/s13229-021-00472-4).
(VI) Atenuación de los síntomas autistas mediante terapia con corticosteroides o inmunoglobulinas terapia;
Una de las características más clásicas de las enfermedades autoinmunes es la respuesta vigorosa al uso terapéutico de las drogas consideradas como los más potentes inmunosupresores y antiinflamatorios: los glucocorticoides (https://pubmed.ncbi.nlm.nih.gov/11457656/). Esto también ocurre en las enfermedades autoinmunes que afectan al sistema nervioso (https://pubmed.ncbi.nlm.nih.gov/11430999/) tales como la esclerosis múltiple (https://pubmed.ncbi.nlm.nih.gov/23229226/). Las principales manifestaciones características (“core features”) del autismo responden de manera similar al tratamiento con corticoides (https://link.springer.com/article/10.1186/1471-2377-14-70; https://pubmed.ncbi.nlm.nih.gov/32168067/; https://www.scielo.br/j/jped/a/PBQNCqJ5L4cyqLXF5hyCxQy/?lang=en#), revelando un mecanismo autoinmune implicado en la fisiopatología del autismo.
De manera similar, las enfermedades autoinmunes pueden responder al tratamiento con inmunoglobulina terapia cuando fallan otros enfoques terapéuticos (https://pubmed.ncbi.nlm.nih.gov/37062358/), como ha sido ampliamente relatado (https://ameripharmaspecialty.com/ivig-and-autoimmune-diseases/). Constituyéndose en evidencia adicional de la participación de mecanismos autoinmunes en el autismo, el mismo efecto ha sido relatado sobre las manifestaciones de este trastorno (https://pubmed.ncbi.nlm.nih.gov/30097568/; https://www.mdpi.com/2075-4426/11/6/488).
(VII) Aparición de trastornos autoinmunes o inmunomediados como comorbilidades en el autismo
Las alergias, el asma, la dermatitis atópica, la rinitis alérgica, la urticaria, la diabetes del tipo 1, la enfermedad inflamatoria intestinal (enfermedad de Crohn) y la psoriasis son comorbilidades del autismo, o sea, son encontradas en portadores de autismo con prevalencia más elevada de que en la población general o no portadora de autismo (https://pubmed.ncbi.nlm.nih.gov/23726766/; https://jlb.onlinelibrary.wiley.com/doi/epdf/10.1189/jlb.1205707; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10619695/; https://pubmed.ncbi.nlm.nih.gov/22511918/; https://www.sciencedirect.com/science/article/abs/pii/S1750946712001018; https://www.scirp.org/journal/paperinformation?paperid=78725; https://pubmed.ncbi.nlm.nih.gov/37939694/). La presencia de dermatitis atópica se asocia no solamente con una mayor probabilidad de autismo, sino también con una mayor gravedad de la condición de autismo (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10619695/), sugiriendo un factor causal común para estas dos condiciones.
(VIII) Alta prevalencia de enfermedades autoinmunes entre parientes consanguíneos (un historial familiar de trastornos autoinmunes aumenta el riesgo de autismo)
En ese grupo de evidencias se demuestra que los niños que tienen familiares portadores de enfermedades autoinmunes, tales como artritis reumatoide, lupus eritematoso sistémico, enfermedad celíaca, colitis ulcerosa, diabetes de tipo 1, hipotiroidismo y/o tiroiditis de Hashimoto, psoriasis y fiebre reumática, tienen más probabilidades de desarrollar autismo
(https://pubmed.ncbi.nlm.nih.gov/19135624/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5373490/; https://pubmed.ncbi.nlm.nih.gov/25981892/; https://jamanetwork.com/journals/jamapediatrics/article-abstract/485932; https://pubmed.ncbi.nlm.nih.gov/14595086/; https://pubmed.ncbi.nlm.nih.gov/10385847/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3115699/; https://pubmed.ncbi.nlm.nih.gov/19581261/; https://pubmed.ncbi.nlm.nih.gov/16598435/; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9025211/).
Los niños cuyos padres son portadores de enfermedades autoinmunes pueden tener un riesgo de un 50% más elevado de desarrollar autismo
(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3115699/).
(IX) Al igual que en las enfermedades autoinmunes clásicas, los beneficios de la administración de colecalciferol también se observan en el autismo;
La “vitamina” D (colecalciferol, cuya forma preactiva es el calcidiol o calcifediol, y cuya forma activa es el calcitriol) tiene una estructura similar a la de las hormonas esteroides (estrógeno, progesterona, testosterona y cortisol). Al igual que las hormonas esteroides, deriva del colesterol, tiene receptores presentes en el núcleo de las células y actúa modificando la actividad genética (modula la actividad de miles de genes (https://www.mdpi.com/2073-4425/14/9/1691; https://www.sciencedirect.com/science/article/pii/S0039128X23000995). Posee receptores nucleares que están presentes en casi todas, si no en todas, las células nucleadas (https://www.sciencedirect.com/science/article/abs/pii/S0003986112001324; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9325172/), así como en las membranas celulares (https://pubmed.ncbi.nlm.nih.gov/17341182/), posee efectos pleiotrópicos (múltiples) sobre el organismo humano (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4045445/; https://www.sciencedirect.com/science/article/abs/pii/B9780323913867000064). Debido a sus múltiples beneficios para la salud humana, el hecho de ser esencialmente producido por la exposición de la piel al sol durante las horas diurnas se considera erróneamente inadecuado (https://www.grassrootshealth.net/blog/shadow-can-tell-right-time-make-vitamin-d/)
y al confinamiento típico de la vida urbana moderna (https://link.springer.com/article/10.1186/s12889-017-4436-z; https://www.sciencedirect.com/science/article/abs/pii/B9780323913867000064), la deficiencia de esta hormona preesteroide (“vitamina” D) ha alcanzado proporciones epidémicas (https://go.gale.com/ps/i.do?id=GALE%7CA592138121&sid=googleScholar&v=2.1&it=r&linkaccess=abs&issn=15228606&p=AONE&sw=w&userGroupName=anon%7E84477f50&aty=open-web-entry; https://academic.oup.com/nutritionreviews/article-abstract/81/10/1290/7071638?redirectedFrom=PDF&login=false; https://www.scielo.br/j/abem/a/78X5HHQSwzZtc435P9CsjCg/?lang=en).
Esta deficiencia tiene profundas consecuencias en el desarrollo cerebral pre y posnatal que pueden extenderse a lo largo de la vida del individuo.
(https://pubs.rsc.org/en/content/articlehtml/2023/fo/d3fo00166k).
Bajos niveles de vitamina D (enmascarados por valores de referencia de laboratorio subdimensionados), (https://www.grassrootshealth.net/wp-content/uploads/2017/05/dip_with_numbers_nmol_051317.pdf; https://www.mdpi.com/2072-6643/16/11/1666), dosis de suplementación “recomendadas” menores a las necesarias (https://www.mdpi.com/2227-9067/1/2/208) y la resistencia genética a sus efectos (https://pubmed.ncbi.nlm.nih.gov/33897704/) ha contribuido para el aumento de la incidencia de una amplia variedad de enfermedades, incluyendo esclerosis múltiple, artritis reumatoide, enfermedades inflamatorias intestinales (enfermedad de Crohn, colitis ulcerosa), enfermedad celíaca, uveítis, enfermedades dermatológicas, espondilitis anquilosante, fibromialgia, diabetes, hipertensión, tuberculosis, Covid-19, y cáncer
(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6032242/; https://link.springer.com/article/10.1007/s00223-019-00577-2; https://www.pnas.org/doi/abs/10.1073/pnas.1200072109; https://www.nature.com/articles/s41430-020-0661-0; https://apcz.umk.pl/QS/article/view/54077; https://www.sciencedirect.com/science/article/abs/pii/S0039625721001685; https://www.nature.com/articles/nrcardio.2009.135; https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.950465/full; https://pubmed.ncbi.nlm.nih.gov/11115787/; https://www.sciencedirect.com/science/article/abs/pii/S0009898114003921; https://www.sciencedirect.com/science/article/abs/pii/S0304395913005411).
Como ya destacaron Wacker y Holick en su publicación de 2012
(https://www.mdpi.com/2072-6643/5/1/111):
“La vitamina D, la vitamina del sol, ha recibido mucha atención recientemente como resultado de un aumento meteórico en el número de publicaciones que muestran que la vitamina D desempeña un papel crucial en una infinidad de funciones fisiológicas y asociando la deficiencia de vitamina D a muchas enfermedades agudas y crónicas, incluidos trastornos del metabolismo del calcio, enfermedades autoinmunes, algunos tipos de cáncer, diabetes mellitus tipo 2, enfermedades cardiovasculares y enfermedades infecciosas”.
Pacientes portadores de enfermedades autoinmunes tratados con altas dosis de vitamina D de forma segura (https://www.mdpi.com/2072-6643/14/8/1575/review_report) pueden tener polimorfismos (SNP) afectando cualquier combinación entre los nueve genes que la vitamina D necesita para producir sus efectos biológicos, incluyendo la regulación del sistema inmune (https://pubmed.ncbi.nlm.nih.gov/33897704/). Evidentemente, esos polimorfismos genéticos pueden provocar resistencia a los efectos de la vitamina D, perjudicando la tolerancia a los autoantígenos (https://pmc.ncbi.nlm.nih.gov/articles/PMC6712894/).
Se destaca el papel fundamental de esta hormona esteroide (en las forma de calcidiol y calcitriol) en la regulación y potencialización del sistema inmune
(https://www.sciencedirect.com/science/article/abs/pii/S1471489210000378; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2861286/; https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.1186635/full), así como en las funciones del SNC (https://academic.oup.com/jbmrplus/article/5/1/e10419/7486306?login=false) tan relevante para esta presentación.
Como destaca Eyles en su artículo (https://www.mdpi.com/2072-6643/5/1/111):
“Ahora también existen, buenas evidencias que asocian la deficiencia de vitamina D gestacional y/o neonatal con un mayor riesgo de trastornos del neurodesarrollo como la esquizofrenia y el autismo, y la deficiencia de vitamina D en adultos a ciertas condiciones degenerativas”.
Tal como se verifica con relación a los beneficios de la vitamina D en enfermedades autoinmunes (https://www.nature.com/articles/ncprheum0855; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2998156/), se verifica también beneficios de la suplementación de vitamina D en el autismo. En su reciente revisión integral (2024), (“umbrella review” – una recopilación y evaluación sistemática de múltiples revisiones sistemáticas y metanálisis sobre un tema de investigación específico,– https://bmjmedicine.bmj.com/content/1/1/e000071), Jiang y colaboradores (https://www.tandfonline.com/doi/full/10.2147/NDS.S470462#abstract) concluyen:
“Basándonos en análisis rigurosos, descubrimos que la deficiencia de vitamina D en el inicio de la vida es un factor de riesgo para el desarrollo del TEA y que la suplementación de vitamina D mejora los principales síntomas del TEA. Nuestro estudio concluye que la suplementación de vitamina D es beneficiosa para individuos con autismo, que la deficiencia de vitamina D en la vida embrionaria temprana aumenta el riesgo de TEA y que nuestro estudio apoya la idea de que la prevención comienza con la suplementación con vitamina D en el inicio de la vida”.
(X) Demostración de una resistencia genética a los efectos inmunorreguladores del colecalciferol (o “vitamina” D – un inhibidor de la actividad Th17) en el autismo.
Tal como se verifica en las enfermedades autoinmunes en general, en las que polimorfismos genéticos relacionados con la actividad de la vitamina D (https://www.cell.com/heliyon/fulltext/S2405-8440(24)03731-9) puede provocar resistencia a los efectos biológicos (incluida la limitación de su papel en la regulación de los sistemas inmune) (https://pubmed.ncbi.nlm.nih.gov/33897704/), también en el autismo se han descrito polimorfismos similares que afectan a los mismos genes (https://www.mdpi.com/2076-3425/7/9/115; https://onlinelibrary.wiley.com/doi/abs/10.1002/aur.2279; https://www.sciencedirect.com/science/article/abs/pii/S0378111916303614; https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6207492/; https://www.sciencedirect.com/science/article/abs/pii/S037837821500119X).
Guerine y colaboradores (https://onlinelibrary.wiley.com/doi/abs/10.1002/aur.2279) observaron inclusive, una correlación entre un polimorfismo (SNP) específico en el gen del receptor de la vitamina D (VDR) y la gravedad del autismo:
“Finalmente, una fuerte asociación gen-dosis del alelo FokI (T) con puntuación más alta en la Escala de Evaluación del Autismo Infantil (Pc = 0,01), y particularmente con el comportamiento de hiperactividad (Pc = 0,006), en niños con el Trastorno del Espectro Autista. “
Esos polimorfismos genéticos, capaces de provocar resistencia a los efectos de la vitamina D, indican que la administración de dosis más elevadas (capaces de compensar el nivel de resistencia), como las utilizadas en la psoriasis y en el vitíligo, puedan restaurar los efectos inmunomoduladores de la vitamina D también en el autismo (https://pubmed.ncbi.nlm.nih.gov/24494059/).
La autoinmunidad presente en el autismo daña el sistema de acceso de la vitamina B9 al Sistema Nervioso Central, produciendo daños secundarios severos a las células nerviosas
El folato (o vitamina B9), cuyas formas naturales son el metilfolato (o metiltetrahidrofolato -la forma activa) y el ácido folínico (o D,L-leucovorina, o 5-formil tetrahidrofolato), es una vitamina del complejo B esencial para el desarrollo y fisiologías normales del SNC. Anormalidades en los niveles de folato ( bajos niveles en el SNC a pesar de los niveles séricos normales) y en las vías relacionadas al folato (polimorfismos genéticos que afectan enzimas involucradas en su metabolismo) fueron identificadas en niños con autismo, lo que caracteriza una condición conocida como síndrome de deficiencia cerebral de folato (“DCF” – también encontrado en la esquizofrenia y en otros trastornos del neurodesarrollo) (https://pmc.ncbi.nlm.nih.gov/articles/PMC8622150/); https://www.degruyter.com/document/doi/10.1515/cclm-2012-0543/html).
El sistema nervioso necesita de concentraciones de folato superiores a las concentraciones sanguíneas. Para alcanzar el tejido nervioso, los folatos necesitan atravesar las barreras que separan la sangre del SNC: las barreras hematoencefálica y hematoliquórica. Por lo tanto, el transporte de folato al SNC está esencialmente mediado por dos sistemas altamente específicos (https://pubmed.ncbi.nlm.nih.gov/32200980/):
(1) Un mecanismo de transporte de alta afinidad (en el que el folato se transfiere para el tejido nervioso con gasto de energía) que es el “receptor alfa de folato” (“folate receptor alpha”); Este es el mecanismo primario de transferencia de folato al cerebro, capaz de bombear folato para niveles tres veces más elevados en la sangre. El transporte se produce a través de un mecanismo llamado potocitosis, donde el receptor unido al folato se internaliza y luego se recicla de nuevo para la membrana de la célula.
(2) Un mecanismo de transporte que permite que los folatos sean transferidos pasivamente para el SNC a través del “transportador de folato reducido” sin utilizar energía. Este sistema secundario sólo puede igualar los niveles de folato en el cerebro con los de la sangre. En otras palabras, las concentraciones de folato en la sangre y en el SNC se mantienen en equilibrio cuando este sistema de transporte es la única opción disponible.
Lo que sería un tercer mecanismo transportador (“acoplado a protones”) en realidad funciona como parte del mecanismo mediado por el receptor de folato alfa a nivel de la barrera hematoliquórica (https://pubmed.ncbi.nlm.nih.gov/19074442/).
Los niños autistas tienen elevados niveles de autoanticuerpos dirigidos contra las neuronas y la concentración de estos anticuerpos se correlaciona directamente con la gravedad de su cuadro neuropsiquiátrico (https://pubmed.ncbi.nlm.nih.gov/22226851/). Varios estudios han demostrado que, entre los niños autistas, el 70% poseen el anticuerpo dirigido contra el receptor de folato (https://pubmed.ncbi.nlm.nih.gov/22230883/; https://pubmed.ncbi.nlm.nih.gov/20668945/).
Dependiendo del tipo de anticuerpo producido, los receptores de folato son bloqueados o destruidos. El primer tipo bloquea la bolsa de unión al folato, mientras que los anticuerpos de unión pueden unirse a otros locales de la estructura proteica del receptor de folato alfa, seguido de la atracción de la cascada del complemento, la activación de citoquinas y finalmente la destrucción del complejo antígeno-anticuerpo (https://pubmed.ncbi.nlm.nih.gov/27068282/). El cerebro depende entonces de la segunda categoría de transportador de folato (transportador de folato reducido) para obtener un nivel mínimo de folato, insuficiente para las necesidades biológicas de las células neuronales. Esa condición (caracterizada por niveles normales de folato en la sangre y reducidos en el líquido cefalorraquídeo o “LCR”) ha sido denominada “síndrome de la deficiencia de folato cerebral” (“Cerebral Folate Deficiency” ou “CFD syndrome”) y también se ha encontrado en otros trastornos neurológicos y neuropsiquiátricos, como el síndrome de Rett, psicosis, esquizofrenia refractaria (intratable), trastornos esquizoafectivos, depresión mayor resistente al tratamiento en adultos, síndrome atáxico espástico y epilepsia intratable entre niños pequeños (https://pubmed.ncbi.nlm.nih.gov/27068282/).
La CFD produce deterioro de las vías metabólicas utilizadas en la síntesis de ácidos nucleicos (y, por lo tanto, inhibición de la neurogénesis – la producción de nuevas neuronas a partir de las células tronco que residen en el SNC). (https://www.sciencedirect.com/science/article/pii/S2405844021021745). Ella también afecta los procesos de metilación (que controlan la expresión de los genes) y las vías de protección contra el efecto destructivo de los llamados radicales libres (un tipo de “desecho celular” que debe ser eliminado continuamente a medida que se forma para que no provoque la destrucción de las células nerviosas) (https://doi.org/10.1016/j.spen.2020.100835).
Una consecuencia importante de la CFD es la reducción de la síntesis de glutatión. El glutatión es un importante antioxidante endógeno que desempeña un papel crucial en la protección de las células contra toxinas exógenas (como metales pesados) y endógenas, particularmente en el SNC (https://pubmed.ncbi.nlm.nih.gov/22528835/). Las anormalidades en el metabolismo del glutatión reducido (GSH) provocan daño oxidativo al ADN celular, las proteínas y los lípidos. Se han documentado anomalías del GSH y marcadores de daño oxidativo en regiones cerebrales post mortem involucradas en el habla, la emoción y el comportamiento social en individuos con autismo (https://doi.org/10.1016/j.spen.2020.100835). De hecho, las anormalidades de metilación y redox son tan frecuentes en el autismo que se ha propuesto que sean utilizados sus biomarcadores en el diagnóstico del TEA (https://doi.org/10.1016/j.spen.2020.100835).
Además, el receptor de folato alfa posee funciones independientes de su papel como transportador de folatos. También desempeña el papel de mantener el repertorio de células tronco que son la fuente de la que constantemente deben originarse nuevas neuronas (neurogénesis) tanto en la vida prenatal como a lo largo de toda la vida del individuo (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5268765/). Su destrucción, por los anticuerpos que se encuentran en niveles elevados en los niños autistas, compromete también la capacidad de recuperación del tejido nervioso dañado por el proceso inflamatorio que caracteriza esta condición.
El síndrome de deficiencia de folato cerebral identificado en el autismo puede ser tratada con altas dosis de ácido folínico por vía oral (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7477301/). Los suplementos de ácido fólico están contraindicados y, si se utilizan, pueden agravar la deficiencia de metilfolato en el LCR (https://pubmed.ncbi.nlm.nih.gov/24494987/; https://pubmed.ncbi.nlm.nih.gov/20668945/). A través de la elevación de los niveles séricos de folato, se obtiene la transferencia de esa vitamina a través de la BHE vía “transportador de folato reducido”. Los niveles séricos elevados (suprafisiológicos) proporcionan entonces la restauración de las concentraciones más elevadas propias (fisiológicas) del SNC.
En el 2013, Frye y sus colegas descubrieron que los anticuerpos dirigidos contra el receptor de alfa folato estaban presentes en el 75% de los niños con autismo cuando se fueran considerados tanto los anticuerpos de bloqueo como los de unión, con el 29% siendo positivos para ambos tipos de anticuerpos (https://pubmed.ncbi.nlm.nih.gov/22230883). La administración de ácido folínico (leucovorina) a una dosis de 2 mg por kg por día (dosis máxima de 50 mg por día) proporcionó mejoras en la comunicación, el lenguaje, la atención y los comportamientos estereotipados en los niños tratados en comparación con los controles portadores de TEA no suplementados. (https://pubmed.ncbi.nlm.nih.gov/22230883). Este enfoque terapéutico fue revisado en el año 2020 (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7477301/), y los autores recomendaron un aumento gradual de la dosis durante las primeras 2 semanas para evitar un posible aumento transitorio de la agitación que ocasionalmente informan los padres. Sin embargo, ellos mismos no observaron tal efecto colateral; verificaron, por el contrario, una mejora de la excitación y de la agitación, después de aproximadamente nueve semanas de tratamiento. Esos resultados indican que, entre los autistas que producen anticuerpos contra el receptor de alfa folato, parte de las manifestaciones autistas pueden ser resultantes o agravadas por el síndrome de la deficiencia de folato cerebral, el que puede corregirse mediante la administración de dosis elevadas de ácido folínico.
La presencia de la variante C677T del gen MTHFR, en particular, constituye un factor de riesgo para el autismo (https://journals.lww.com/psychgenetics/abstract/2009/08000/aberrations_in_folate_metabolic_pathway_and.2.aspx) y puede interactuar con polimorfismos que afectan otros genes relacionados con el metabolismo del folato [como el polimorfismo de deleción de 19 pares de bases de la enzima dihidrofolato reductasa (DHFR) y el SNP G80A que afecta el gen del transportador de folato reducido] para aumentar el riesgo de autismo (https://pubmed.ncbi.nlm.nih.gov/17597297/). Evidentemente, estos polimorfismos pueden exacerbar el síndrome de deficiencia cerebral de folato, particularmente cuando ocurren en asociación. La identificación de un polimorfismo que afecta al gen DHFR como otro factor de riesgo para el autismo (https://pubmed.ncbi.nlm.nih.gov/17597297/) enfatiza aún más la necesidad de evitar la suplementación de ácido fólico en niños autistas, ya que el metabolismo (reducción) de ese folato sintético requiere la actividad de la DHFR.
MTHFR requiere vitamina B2 (riboflavina), que actúa como su cofactor (https://www.ncbi.nlm.nih.gov/books/NBK6145/). A su vez, la deficiencia de riboflavina puede deberse a factores genéticos prevalentes en la población en general (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1918332/), potencialmente representando un factor adicional que contribuye para la susceptibilidad al autismo. Por tanto, la administración de dosis suprafisiológicas de riboflavina puede resultar beneficiosa.
Concluyendo este segundo Editorial, las evidencias acumuladas durante las últimas décadas deja clara la participación fundamental de los mecanismos autoinmunes en la fisiopatología de la encefalitis crónica asociada al autismo (caracterizada en el primer Editorial). La tríada considerada “la marca registrada” de una enfermedad autoinmune (inflamación, producción de autoanticuerpos e infiltración linfocitaria) está presente en el autismo. En el próximo Editorial veremos que la afirmación de que “el origen de la autoinmunidad en el autismo es desconocida” (https://pubmed.ncbi.nlm.nih.gov/22226851/ – página 465) ya no se sustenta.
Leia as postagens pelo índice mensal:
- outubro 2025
- julho 2025
- junho 2025
- abril 2025
- março 2025
- fevereiro 2025
- janeiro 2025
- dezembro 2024
- novembro 2024
- outubro 2024
- setembro 2024
- agosto 2024
- julho 2024
- julho 2021
- maio 2021
- abril 2021
- fevereiro 2021
- janeiro 2021
- dezembro 2020
- novembro 2020
- outubro 2020
- setembro 2020
- agosto 2020
- julho 2020
- junho 2020
- maio 2020
- abril 2020
Leia as últimas postagens:
- Evidencias científicas de que las vacunas actualmente usadas mundialmente causan Trastorno del Espectro Autista (TEA), un “Síndrome Autoinmune (Auto-inflamatorio) inducido por Adyuvantes” (ASIA). El mecanismo del “caballo de Troya”. Este es el tercer editorial de este sitio sobre la causa del autismo
 Primera editorial: https://wp.me/pbW3AH-1H7 Segundo editorial: https://wp.me/pbW3AH-1IX Traducido al español por Carmen Sanabria Ayala Índice 1) Introducción 2) Negacionismo 3) El impacto del autismo en el sistema educativo 4) Identificando los componentes de las vacunas como la causa raíz del TEA y el aumento del calendario de vacunación, incluidas las vacunaciones múltiples simultáneas y recurrentes como… Leia mais: Evidencias científicas de que las vacunas actualmente usadas mundialmente causan Trastorno del Espectro Autista (TEA), un “Síndrome Autoinmune (Auto-inflamatorio) inducido por Adyuvantes” (ASIA). El mecanismo del “caballo de Troya”. Este es el tercer editorial de este sitio sobre la causa del autismo
Primera editorial: https://wp.me/pbW3AH-1H7 Segundo editorial: https://wp.me/pbW3AH-1IX Traducido al español por Carmen Sanabria Ayala Índice 1) Introducción 2) Negacionismo 3) El impacto del autismo en el sistema educativo 4) Identificando los componentes de las vacunas como la causa raíz del TEA y el aumento del calendario de vacunación, incluidas las vacunaciones múltiples simultáneas y recurrentes como… Leia mais: Evidencias científicas de que las vacunas actualmente usadas mundialmente causan Trastorno del Espectro Autista (TEA), un “Síndrome Autoinmune (Auto-inflamatorio) inducido por Adyuvantes” (ASIA). El mecanismo del “caballo de Troya”. Este es el tercer editorial de este sitio sobre la causa del autismo - Norwegian version of the third editorial on the causes of autism
 Translation, Dr. Geir Flatabø Vitenskapelige bevis / evidens for at vaksiner i dag brukt over hele verden forårsaker Autisme Spekter Forstyrrelse (ASD) – en “Autoimmun (Auto Inflammatorisk) Syndrom Indusert av Adjuvans” (ASIA). “Trojansk hest” -mekanisme. “Stille infeksjon” i hjernevevet. Dette er den tredje redaksjonelle artikkel på dette nettstedet om autisme. REGISTER / INNHOLD 1) Innledning2)… Leia mais: Norwegian version of the third editorial on the causes of autism
Translation, Dr. Geir Flatabø Vitenskapelige bevis / evidens for at vaksiner i dag brukt over hele verden forårsaker Autisme Spekter Forstyrrelse (ASD) – en “Autoimmun (Auto Inflammatorisk) Syndrom Indusert av Adjuvans” (ASIA). “Trojansk hest” -mekanisme. “Stille infeksjon” i hjernevevet. Dette er den tredje redaksjonelle artikkel på dette nettstedet om autisme. REGISTER / INNHOLD 1) Innledning2)… Leia mais: Norwegian version of the third editorial on the causes of autism - Romanian version – Dovezi științifice că vaccinurile utilizate în prezent la nivel mondial cauzează tulburări din spectrul autist (TSA)
 Translated into romanian by Dr. Dominique Heidenfelder-Ambrosio Dovezi științifice că vaccinurile utilizate în prezent la nivel mondial cauzează tulburări din spectrul autist (TSA) – un „sindrom autoimun (autoinflamator) indus de adjuvanți” (ASIA). Mecanismul „calului troian”. „Infecția silențioasă” a țesutului cerebral. Acesta este al treilea editorial de pe acest site web despre cauza autismului. Index 1)… Leia mais: Romanian version – Dovezi științifice că vaccinurile utilizate în prezent la nivel mondial cauzează tulburări din spectrul autist (TSA)
Translated into romanian by Dr. Dominique Heidenfelder-Ambrosio Dovezi științifice că vaccinurile utilizate în prezent la nivel mondial cauzează tulburări din spectrul autist (TSA) – un „sindrom autoimun (autoinflamator) indus de adjuvanți” (ASIA). Mecanismul „calului troian”. „Infecția silențioasă” a țesutului cerebral. Acesta este al treilea editorial de pe acest site web despre cauza autismului. Index 1)… Leia mais: Romanian version – Dovezi științifice că vaccinurile utilizate în prezent la nivel mondial cauzează tulburări din spectrul autist (TSA) - Wissenschaftliche Erkenntnisse belegen, dass die derzeit weltweit verwendeten Impfstoffe Autismus Spektrum-Störungen verursachen
 Translated into German by Dr. Dominique Heidenfelder-Ambrosio Störungen (ASS) verursachen – ein durch Adjuvantien induziertes Autoimmunsyndrom (ASIA). Der „Trojanische Pferd“-Mechanismus. Eine „stille Infektion“ des Hirngewebes. Dies ist der dritte Leitartikel auf dieser Website zu den Ursachen von Autismus Index 1) Einleitung 2) Leugnung 3) Die Auswirkungen von Autismus auf das Bildungssystem 4) Identifizierung von Impfstoffbestandteilen… Leia mais: Wissenschaftliche Erkenntnisse belegen, dass die derzeit weltweit verwendeten Impfstoffe Autismus Spektrum-Störungen verursachen
Translated into German by Dr. Dominique Heidenfelder-Ambrosio Störungen (ASS) verursachen – ein durch Adjuvantien induziertes Autoimmunsyndrom (ASIA). Der „Trojanische Pferd“-Mechanismus. Eine „stille Infektion“ des Hirngewebes. Dies ist der dritte Leitartikel auf dieser Website zu den Ursachen von Autismus Index 1) Einleitung 2) Leugnung 3) Die Auswirkungen von Autismus auf das Bildungssystem 4) Identifizierung von Impfstoffbestandteilen… Leia mais: Wissenschaftliche Erkenntnisse belegen, dass die derzeit weltweit verwendeten Impfstoffe Autismus Spektrum-Störungen verursachen - Prove scientifiche che i vaccini attualmente utilizzati in tutto il mondo causano Disturbi dello Spettro Autistico
 Translated into Italian by Dr. Dominique Heidenfelder-Ambrosio “Sindrome Autoimmune (Auto-infiammatoria) indotta da Adiuvanti” (ASIA). Il meccanismo del “cavallo di Troia”. “Infezione silenziosa” del tessuto cerebrale. Questo è il terzo editoriale su questo sito web sulla causa dell’autismo Indice 1) Introduzione 2) Negazionismo 3) L’impatto dell’autismo sul sistema educativo 4) Identificare i componenti del vaccino come… Leia mais: Prove scientifiche che i vaccini attualmente utilizzati in tutto il mondo causano Disturbi dello Spettro Autistico
Translated into Italian by Dr. Dominique Heidenfelder-Ambrosio “Sindrome Autoimmune (Auto-infiammatoria) indotta da Adiuvanti” (ASIA). Il meccanismo del “cavallo di Troia”. “Infezione silenziosa” del tessuto cerebrale. Questo è il terzo editoriale su questo sito web sulla causa dell’autismo Indice 1) Introduzione 2) Negazionismo 3) L’impatto dell’autismo sul sistema educativo 4) Identificare i componenti del vaccino come… Leia mais: Prove scientifiche che i vaccini attualmente utilizzati in tutto il mondo causano Disturbi dello Spettro Autistico - Second editorial on the causes of autism, translated into norwegian
 Translated by Geir Flatabø En kronisk inflammatorisk prosess skader hjernen til barn som får en autisme diagnose, denne har en autoimmun komponent. En av konsekvensene er at transport av vitamin B9 (Folinsyre) til nervesystemet hemmes. Dette medfører alvorlig redusert funksjon i stoffskiftet som bruker vit B9, og selv om de er godt dokumentert, har kunnskapen… Leia mais: Second editorial on the causes of autism, translated into norwegian
Translated by Geir Flatabø En kronisk inflammatorisk prosess skader hjernen til barn som får en autisme diagnose, denne har en autoimmun komponent. En av konsekvensene er at transport av vitamin B9 (Folinsyre) til nervesystemet hemmes. Dette medfører alvorlig redusert funksjon i stoffskiftet som bruker vit B9, og selv om de er godt dokumentert, har kunnskapen… Leia mais: Second editorial on the causes of autism, translated into norwegian - Evidências científicas de que as vacinas atualmente usadas mundialmente causam Transtorno do Espectro Autista – Este é o terceiro editorial neste site sobre a causa do autismo
 Evidências científicas de que as vacinas atualmente usadas mundialmente causam Transtorno do Espectro Autista (TEA) – uma “Síndrome Autoimune (Auto-inflamatória) induzida por Adjuvantes” (ASIA). O mecanismo do “cavalo de Tróia”. Este é o terceiro editorial neste site sobre a causa do autismo. Índice 1) Introdução 2) Negacionismo 3) O impacto do autismo no sistema educacional… Leia mais: Evidências científicas de que as vacinas atualmente usadas mundialmente causam Transtorno do Espectro Autista – Este é o terceiro editorial neste site sobre a causa do autismo
Evidências científicas de que as vacinas atualmente usadas mundialmente causam Transtorno do Espectro Autista (TEA) – uma “Síndrome Autoimune (Auto-inflamatória) induzida por Adjuvantes” (ASIA). O mecanismo do “cavalo de Tróia”. Este é o terceiro editorial neste site sobre a causa do autismo. Índice 1) Introdução 2) Negacionismo 3) O impacto do autismo no sistema educacional… Leia mais: Evidências científicas de que as vacinas atualmente usadas mundialmente causam Transtorno do Espectro Autista – Este é o terceiro editorial neste site sobre a causa do autismo - Scientific evidence that vaccines currently used worldwide cause Autism Spectrum Disorder – Third editorial on this website on the cause of autism(Scientific evidence that vaccines currently used worldwide cause Autism Spectrum Disorder (ASD) – an “Autoimmune (Auto-inflammatory) Syndrome induced by Adjuvants” (ASIA). The “Trojan horse” mechanism. “Silent infection” of the brain tissue.) Index 1) Introduction 2) Denialism 3) The impact of autism on the education system 4) Identifying vaccine components as the root cause of ASD and… Leia mais: Scientific evidence that vaccines currently used worldwide cause Autism Spectrum Disorder – Third editorial on this website on the cause of autism
- Le processus inflammatoire chronique qui endommage le cerveau des enfants autistes présente une composante auto-immune incontestable – deuxième éditorial sur ce site concernant les causes de l’autisme
 Le processus inflammatoire chronique qui endommage le cerveau des enfants autistes présente une composante auto-immune incontestable. Parmi les conséquences, le transport de la vitamine B9 (folate) vers le système nerveux peut être inhibé, entraînant des implications thérapeutiques profondes qui, bien que documentées, ont été largement ignorées. Il s’agit du deuxième éditorial sur ce site concernant… Leia mais: Le processus inflammatoire chronique qui endommage le cerveau des enfants autistes présente une composante auto-immune incontestable – deuxième éditorial sur ce site concernant les causes de l’autisme
Le processus inflammatoire chronique qui endommage le cerveau des enfants autistes présente une composante auto-immune incontestable. Parmi les conséquences, le transport de la vitamine B9 (folate) vers le système nerveux peut être inhibé, entraînant des implications thérapeutiques profondes qui, bien que documentées, ont été largement ignorées. Il s’agit du deuxième éditorial sur ce site concernant… Leia mais: Le processus inflammatoire chronique qui endommage le cerveau des enfants autistes présente une composante auto-immune incontestable – deuxième éditorial sur ce site concernant les causes de l’autisme - Indicação de livro: Além da Vitamina D, versão em português
 Histórias emocionantes mostram como superar doenças autoimunes Mais do que um livro sobre saúde, este primeiro volume da série “Além da Vitamina D” relata histórias de pacientes, seus familiares, médicos, e médicos que são também pacientes do tratamento que devolve qualidade de vida a portadores de inúmeras doenças consideradas incuráveis. Enquanto informa, ao longo de… Leia mais: Indicação de livro: Além da Vitamina D, versão em português
Histórias emocionantes mostram como superar doenças autoimunes Mais do que um livro sobre saúde, este primeiro volume da série “Além da Vitamina D” relata histórias de pacientes, seus familiares, médicos, e médicos que são também pacientes do tratamento que devolve qualidade de vida a portadores de inúmeras doenças consideradas incuráveis. Enquanto informa, ao longo de… Leia mais: Indicação de livro: Além da Vitamina D, versão em português - Saiba as consequências da deficiência de Vitamina D – Dra. Satya Nahu
 Leia as postagens pelo índice mensal: Leia as últimas postagens:
Leia as postagens pelo índice mensal: Leia as últimas postagens: - Deficiência de vitamina D atende aos critérios de Hill para causalidade na suscetibilidade, complicações e mortalidade do SARS-CoV-2: uma revisão sistemática
 Fonte: https://www.mdpi.com/2072-6643/17/3/599 Os ensaios clínicos demonstram consistentemente uma correlação inversa entre os níveis séricos de 25-hidroxivitamina D [25(OH)D; calcifediol] e o risco de doença sintomática por SARS-CoV-2, complicações e mortalidade. Esta revisão sistemática (RS), guiada pelos critérios de causalidade de Bradford Hill, analisou 294 manuscritos revisados por pares publicados entre dezembro de 2019 e novembro… Leia mais: Deficiência de vitamina D atende aos critérios de Hill para causalidade na suscetibilidade, complicações e mortalidade do SARS-CoV-2: uma revisão sistemática
Fonte: https://www.mdpi.com/2072-6643/17/3/599 Os ensaios clínicos demonstram consistentemente uma correlação inversa entre os níveis séricos de 25-hidroxivitamina D [25(OH)D; calcifediol] e o risco de doença sintomática por SARS-CoV-2, complicações e mortalidade. Esta revisão sistemática (RS), guiada pelos critérios de causalidade de Bradford Hill, analisou 294 manuscritos revisados por pares publicados entre dezembro de 2019 e novembro… Leia mais: Deficiência de vitamina D atende aos critérios de Hill para causalidade na suscetibilidade, complicações e mortalidade do SARS-CoV-2: uma revisão sistemática - Norwegian version. Autisme hjernen er påvirket av en kronisk inflammatorisk prosess som kan og bør behandles. Dette er den første redaksjonelle artikkelen på denne nettsiden om årsakene til autismeTranslated by Geir Flatabø Autisme hjernen er påvirket av en kronisk inflammatorisk prosess som kan og bør behandles. Dette er den første redaksjonelle artikkelen på denne nettsiden om årsakene til autisme Autisme ble først beskrevet av Leo Kanner ( en barnepsykiater ved Johns Hopkins School of Medicine ) i 1943. Han kalte det “autistiske lidelser”… Leia mais: Norwegian version. Autisme hjernen er påvirket av en kronisk inflammatorisk prosess som kan og bør behandles. Dette er den første redaksjonelle artikkelen på denne nettsiden om årsakene til autisme
- “A Grande Indústria Farmacêutica é dona do seu médico”
 “A Big Pharma é dona do seu médico: “A coisa toda é bananas”. “Nossos médicos estudaram medicina em escolas financiadas pela Big Pharma. “A referência dos médicos científicos sobre a segurança e eficácia de vacinas e outros medicamentos é financiada pela Big Pharma. “As agências reguladoras de saúde que deveriam manter a Big Pharma sob… Leia mais: “A Grande Indústria Farmacêutica é dona do seu médico”
“A Big Pharma é dona do seu médico: “A coisa toda é bananas”. “Nossos médicos estudaram medicina em escolas financiadas pela Big Pharma. “A referência dos médicos científicos sobre a segurança e eficácia de vacinas e outros medicamentos é financiada pela Big Pharma. “As agências reguladoras de saúde que deveriam manter a Big Pharma sob… Leia mais: “A Grande Indústria Farmacêutica é dona do seu médico” - Como a vitamina D mudou a vida de uma médica com esclerose múltipla – sua jornada e uma entrevista -Protocolo CoimbraFonte: https://www.grassrootshealth.net/blog/vitamin-d-changed-life-doctor-ms-journey-interview/ “O Protocolo de Coimbra ajudou milhares de pessoas em todo o mundo a superar as suas doenças auto-imunes usando doses extremamente elevadas de vitamina D. Não se trate sem a orientação de um profissional qualificado.” “Narrativa de Satya L. Nahu, MD; não deixe de ler a curta entrevista abaixo também! “Minha história com… Leia mais: Como a vitamina D mudou a vida de uma médica com esclerose múltipla – sua jornada e uma entrevista -Protocolo Coimbra
- Dr. Toby Rogers: a aprovação do uso de alumínio em vacinas foi baseado em um estudo de apenas quatro coelhos
 “O FDA e o CDC aprovaram o alumínio como ‘seguro e eficaz’ em vacinas, com base em um estudo de apenas 4 coelhos.” Dr. Toby Rogers, PhD “Cada injeção no cronograma infantil do CDC, de acordo com o melhor conjunto de dados de vacinas do mundo, causa mais danos do que benefícios.” “Os adjuvantes de… Leia mais: Dr. Toby Rogers: a aprovação do uso de alumínio em vacinas foi baseado em um estudo de apenas quatro coelhos
“O FDA e o CDC aprovaram o alumínio como ‘seguro e eficaz’ em vacinas, com base em um estudo de apenas 4 coelhos.” Dr. Toby Rogers, PhD “Cada injeção no cronograma infantil do CDC, de acordo com o melhor conjunto de dados de vacinas do mundo, causa mais danos do que benefícios.” “Os adjuvantes de… Leia mais: Dr. Toby Rogers: a aprovação do uso de alumínio em vacinas foi baseado em um estudo de apenas quatro coelhos - Robert Kennedy Jr. acusa Congresso dos EUA de receber dinheiro da indústria farmacêutica em sua sabatina no Senado, no dia 29 de janeiro último – vídeo com legendas em português
 *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens: (Continua)
*Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens: (Continua) - Estudo de “de cair o queixo” descobre que crianças vacinadas têm 170% maior risco de autismo
 Fonte:‘Jaw-dropping’ Study Finds Vaccinated Children Have 170% Higher Risk of Autism “O estudo revisado por pares também descobriu que as crianças vacinadas tinham uma probabilidade 212% maior de desenvolver outros distúrbios do neurodesenvolvimento, incluindo TDAH, epilepsia / apreensão, inflamação cerebral e distúrbios de tic e aprendizagem. As crianças vacinadas têm uma chance 170% maior de… Leia mais: Estudo de “de cair o queixo” descobre que crianças vacinadas têm 170% maior risco de autismo
Fonte:‘Jaw-dropping’ Study Finds Vaccinated Children Have 170% Higher Risk of Autism “O estudo revisado por pares também descobriu que as crianças vacinadas tinham uma probabilidade 212% maior de desenvolver outros distúrbios do neurodesenvolvimento, incluindo TDAH, epilepsia / apreensão, inflamação cerebral e distúrbios de tic e aprendizagem. As crianças vacinadas têm uma chance 170% maior de… Leia mais: Estudo de “de cair o queixo” descobre que crianças vacinadas têm 170% maior risco de autismo - Como trabalha o seu cérebro para a saúde ou para a doença – vídeo com legendas
 *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens:
*Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens: - Anthony Fauci, um dos responsáveis pela “pandemia”, e pelo uso de vacinas como arma de guerra biológica, recebeu perdão antecipado nos últimos momentos do governo Biden – vídeo em português
 *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens:
*Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Links para as últimas 100 postagens: - Vacinação e distúrbios do neurodesenvolvimento: um estudo de crianças de nove anos matriculadas no Medicaid – Transtorno do espectro autista. Publicação de 23.01.2025 na Science, Public Health Policy and the Law
 As taxas de distúrbios do neurodesenvolvimento (NDD) aumentaram mais de dez vezes durante a década de 1980 e uma em cada seis crianças dos EUA foi diagnosticada com uma deficiência de desenvolvimento entre 2009 e 2017. NDD é distúrbio do neurodesenvolvimento Leia na fonte da publicação: Vaccination and Neurodevelopmental Disorders: A Study of Nine-Year-Old Children… Leia mais: Vacinação e distúrbios do neurodesenvolvimento: um estudo de crianças de nove anos matriculadas no Medicaid – Transtorno do espectro autista. Publicação de 23.01.2025 na Science, Public Health Policy and the Law
As taxas de distúrbios do neurodesenvolvimento (NDD) aumentaram mais de dez vezes durante a década de 1980 e uma em cada seis crianças dos EUA foi diagnosticada com uma deficiência de desenvolvimento entre 2009 e 2017. NDD é distúrbio do neurodesenvolvimento Leia na fonte da publicação: Vaccination and Neurodevelopmental Disorders: A Study of Nine-Year-Old Children… Leia mais: Vacinação e distúrbios do neurodesenvolvimento: um estudo de crianças de nove anos matriculadas no Medicaid – Transtorno do espectro autista. Publicação de 23.01.2025 na Science, Public Health Policy and the Law - Múltiplas vacinações simultâneas estão associadas à probabilidade de hospitalização ou morte por uma reação adversa
 (Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com) Múltiplas vacinações simultâneas (comumente quatro a oito injeções) são aplicadas de forma recorrente a intervalos tão curtos quanto dois meses ao longo dos primeiros anos de vida (https://www.cdc.gov/vaccines/hcp/imz-schedules/child-adolescent-age.html). Nenhuma dessas vacinas é objeto de… Leia mais: Múltiplas vacinações simultâneas estão associadas à probabilidade de hospitalização ou morte por uma reação adversa
(Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com) Múltiplas vacinações simultâneas (comumente quatro a oito injeções) são aplicadas de forma recorrente a intervalos tão curtos quanto dois meses ao longo dos primeiros anos de vida (https://www.cdc.gov/vaccines/hcp/imz-schedules/child-adolescent-age.html). Nenhuma dessas vacinas é objeto de… Leia mais: Múltiplas vacinações simultâneas estão associadas à probabilidade de hospitalização ou morte por uma reação adversa - Reconsideração dos níveis de dose segura pediátrica imunoterapêutica de alumínio – Reconsideration of the immunotherapeutic pediatric safe dose levels of aluminum
 Fonte: https://www.sciencedirect.com/science/article/pii/S0946672X17300950#bib0210 Conclusões do artigo: “Os níveis de alumínio nas vacinas são baseados na eficácia imunológica e ignoram o cálculo da segurança conforme o peso corporal.” “Vários erros críticos foram cometidos na avaliação da dosagem pediátrica de alumínio em vacinas.” “As inferências de segurança das doses de de alumínio em vacinas se basearam apenas em… Leia mais: Reconsideração dos níveis de dose segura pediátrica imunoterapêutica de alumínio – Reconsideration of the immunotherapeutic pediatric safe dose levels of aluminum
Fonte: https://www.sciencedirect.com/science/article/pii/S0946672X17300950#bib0210 Conclusões do artigo: “Os níveis de alumínio nas vacinas são baseados na eficácia imunológica e ignoram o cálculo da segurança conforme o peso corporal.” “Vários erros críticos foram cometidos na avaliação da dosagem pediátrica de alumínio em vacinas.” “As inferências de segurança das doses de de alumínio em vacinas se basearam apenas em… Leia mais: Reconsideração dos níveis de dose segura pediátrica imunoterapêutica de alumínio – Reconsideration of the immunotherapeutic pediatric safe dose levels of aluminum - Alumínio no tecido cerebral no autismo – Aluminium in brain tissue in autism – ScienceDirect
 Fonte: https://www.sciencedirect.com/science/article/pii/S0946672X17308763 “…o fato de termos encontrado alumínio em cada amostra de tecido cerebral, congelada ou fixada, sugere fortemente que os indivíduos com diagnóstico de Transtorno do Espectro Autista têm níveis extraordinariamente altos de alumínio em seu tecido cerebral e que este alumínio está particularmente associado a células não neuronais incluindo micróglia e outras células… Leia mais: Alumínio no tecido cerebral no autismo – Aluminium in brain tissue in autism – ScienceDirect
Fonte: https://www.sciencedirect.com/science/article/pii/S0946672X17308763 “…o fato de termos encontrado alumínio em cada amostra de tecido cerebral, congelada ou fixada, sugere fortemente que os indivíduos com diagnóstico de Transtorno do Espectro Autista têm níveis extraordinariamente altos de alumínio em seu tecido cerebral e que este alumínio está particularmente associado a células não neuronais incluindo micróglia e outras células… Leia mais: Alumínio no tecido cerebral no autismo – Aluminium in brain tissue in autism – ScienceDirect - Autystyczny mózg jest uszkodzony przez nieleczony przewlekły proces zapalny. Pierwsza publikacja na tej stronie o przyczynach autyzmu
 Translated into Polish by Father Jacek Maria Norkowski OP, Md. PhD., former teacher of moral theology in Pontifical University of Saint Thomas Aquinas in Rome. Autyzm został po raz pierwszy opisany przez Leo Kannera (Psychiatra dziecięcy w Johns Hopkins School of Medicine) w 1943 roku, który nazwał to “zaburzeniami autystycznymi”(https://www.autismtruths.org/pdf/autistic%20disturbances%20of%20affective%20contact%20-%20leo%20kanner.pdf). Dziewieć lat później, dolegliwość, jaką… Leia mais: Autystyczny mózg jest uszkodzony przez nieleczony przewlekły proces zapalny. Pierwsza publikacja na tej stronie o przyczynach autyzmu
Translated into Polish by Father Jacek Maria Norkowski OP, Md. PhD., former teacher of moral theology in Pontifical University of Saint Thomas Aquinas in Rome. Autyzm został po raz pierwszy opisany przez Leo Kannera (Psychiatra dziecięcy w Johns Hopkins School of Medicine) w 1943 roku, który nazwał to “zaburzeniami autystycznymi”(https://www.autismtruths.org/pdf/autistic%20disturbances%20of%20affective%20contact%20-%20leo%20kanner.pdf). Dziewieć lat później, dolegliwość, jaką… Leia mais: Autystyczny mózg jest uszkodzony przez nieleczony przewlekły proces zapalny. Pierwsza publikacja na tej stronie o przyczynach autyzmu - Der chronische Entzündungsprozess, der das Gehirn autistischer Kinder betrifft, hat unbestreitbar eine Autoimmunkomponente – Zweiter Leitartikel auf dieser Website zu den Ursachen von AutismusBewertet von:Dr. Melanie WeissWilhelmstraße 17342489 Wülfrathinfo@neurologie-wuelfrath.de (Erster ins Deutsche übersetzter Leitartikel: https://wp.me/pbW3AH-1Gu) Der chronische Entzündungsprozess, der das Gehirn autistischer Kinder schädigt, weist eindeutig eine autoimmune Komponente auf. Infolgedessen wird der Transport von Vitamin B9 (Folat) zum Nervensystem gehemmt, was zu tiefgreifenden Auswirkungen führen kann, die zwar dokumentiert, aber weitgehend ignoriert wurden. Dies ist der zweite… Leia mais: Der chronische Entzündungsprozess, der das Gehirn autistischer Kinder betrifft, hat unbestreitbar eine Autoimmunkomponente – Zweiter Leitartikel auf dieser Website zu den Ursachen von Autismus
- O papel da vitamina D nos resultados dos cuidados intensivos em pacientes com COVID-19
 Fonte: O papel da vitamina D nos resultados dos cuidados intensivos em pacientes com COVID-19: evidência de uma meta-análise guarda-chuva de estudos intervencionistas e observacionais Publicado online pela Cambridge University Press: 24 de Abril de 2024 Conclusão do estudo “No geral, o estado da vitamina D é um fator crítico que influencia a taxa de… Leia mais: O papel da vitamina D nos resultados dos cuidados intensivos em pacientes com COVID-19
Fonte: O papel da vitamina D nos resultados dos cuidados intensivos em pacientes com COVID-19: evidência de uma meta-análise guarda-chuva de estudos intervencionistas e observacionais Publicado online pela Cambridge University Press: 24 de Abril de 2024 Conclusão do estudo “No geral, o estado da vitamina D é um fator crítico que influencia a taxa de… Leia mais: O papel da vitamina D nos resultados dos cuidados intensivos em pacientes com COVID-19 - Robert Kennedy Jr denunciou que a indústria farmacêutica sabia que suas vacinas causavam autismo – vídeo com legendas
 De acordo com RFK Jr., uma denunciante lhe forneceu uma transcrição de uma reunião secreta envolvendo autoridades do FDA, CDC, NIH e OMS, provando que as autoridades de saúde pública sabiam que as vacinas infantis estavam causando autismo. “Eles observaram a vacina contra hepatite B administrada durante os primeiros 30 dias e compararam com crianças… Leia mais: Robert Kennedy Jr denunciou que a indústria farmacêutica sabia que suas vacinas causavam autismo – vídeo com legendas
De acordo com RFK Jr., uma denunciante lhe forneceu uma transcrição de uma reunião secreta envolvendo autoridades do FDA, CDC, NIH e OMS, provando que as autoridades de saúde pública sabiam que as vacinas infantis estavam causando autismo. “Eles observaram a vacina contra hepatite B administrada durante os primeiros 30 dias e compararam com crianças… Leia mais: Robert Kennedy Jr denunciou que a indústria farmacêutica sabia que suas vacinas causavam autismo – vídeo com legendas - Os adjuvantes de vacinas de alumínio contribuem para o aumento da prevalência do autismo?
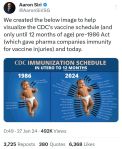 “os recém-nascidos recebem uma dose de alumínio equivalente a 10 (dez) vacinas contra hepatite B administradas simultaneamente a um adulto de 70 kg” Tabela número 5, extraída da publicação “Do aluminum vaccine adjuvants contribute to the rising prevalence of autism? (Os adjuvantes* de vacinas de alumínio contribuem para o aumento da prevalência do autismo?)”: https://www.sciencedirect.com/science/article/abs/pii/S0162013411002212… Leia mais: Os adjuvantes de vacinas de alumínio contribuem para o aumento da prevalência do autismo?
“os recém-nascidos recebem uma dose de alumínio equivalente a 10 (dez) vacinas contra hepatite B administradas simultaneamente a um adulto de 70 kg” Tabela número 5, extraída da publicação “Do aluminum vaccine adjuvants contribute to the rising prevalence of autism? (Os adjuvantes* de vacinas de alumínio contribuem para o aumento da prevalência do autismo?)”: https://www.sciencedirect.com/science/article/abs/pii/S0162013411002212… Leia mais: Os adjuvantes de vacinas de alumínio contribuem para o aumento da prevalência do autismo? - Il processo infiammatorio cronico che sta vivendo il cervello dei bambini autistici ha un’innegabile componente autoimmune – Secondo Editoriale di questo Sito sulle cause dell’autismoTraduzione fatta da Dottoressa Francesca Micheluccifmichelucci22@gmail.com, Pisa Primo editoriale tradotto in italiano https://wp.me/pbW3AH-1FM Il processo infiammatorio cronico che danneggia il cervello dei bambini autistici presenta un’innegabile componente autoimmune. Tra le varie conseguenze, può essere inibito il trasporto della vitamina B9 ( folato ) attraverso il sistema nervoso , portando a profonde implicazioni terapeutiche che, sebbene… Leia mais: Il processo infiammatorio cronico che sta vivendo il cervello dei bambini autistici ha un’innegabile componente autoimmune – Secondo Editoriale di questo Sito sulle cause dell’autismo
- As chantagens da Pfizer para impor sua vacina aos países durante a pandemia – Pfizer’s blackmail to impose its vaccine on countries during the pandemic – com legendas
 Leia as postagens pelo índice mensal: Leia as últimas postagens:
Leia as postagens pelo índice mensal: Leia as últimas postagens: - Segundo Editorial sobre las Causas de Autismo, traducido al español. El proceso inflamatorio crónico que está dañando el cerebro de los niños autistas tiene un innegable componente autoinmune.(Primera editorial: https://wp.me/pbW3AH-1H7) Traducido al español por Carmen Sanabria Ayala Entre las consecuencias está la inhibición del transporte de vitamina B9 (folato) al sistema nervioso, con profundas implicaciones terapéuticas que, a pesar de estar documentadas, han sido en gran medida ignoradas. Segundo Editorial de este Site sobre las causas del autismo. Fundamentándose en la lógica… Leia mais: Segundo Editorial sobre las Causas de Autismo, traducido al español. El proceso inflamatorio crónico que está dañando el cerebro de los niños autistas tiene un innegable componente autoinmune.
- Uma associação positiva encontrada entre a prevalência do autismo e a aceitação da vacinação infantil em toda a população dos EUA
 Fonte: https://pubmed.ncbi.nlm.nih.gov/21623535/ NIH, National Library of Medicine, “Um site oficial do governo dos Estados Unidos” Publicação de 2011 PMID: Endereço de 21623535DOI: 10.1080/15287394.2011.573736 “Abstrato “A razão para o rápido aumento do autismo nos Estados Unidos que começou na década de 1990 é um mistério. Embora os indivíduos provavelmente tenham uma predisposição genética para desenvolver autismo,… Leia mais: Uma associação positiva encontrada entre a prevalência do autismo e a aceitação da vacinação infantil em toda a população dos EUA
Fonte: https://pubmed.ncbi.nlm.nih.gov/21623535/ NIH, National Library of Medicine, “Um site oficial do governo dos Estados Unidos” Publicação de 2011 PMID: Endereço de 21623535DOI: 10.1080/15287394.2011.573736 “Abstrato “A razão para o rápido aumento do autismo nos Estados Unidos que começou na década de 1990 é um mistério. Embora os indivíduos provavelmente tenham uma predisposição genética para desenvolver autismo,… Leia mais: Uma associação positiva encontrada entre a prevalência do autismo e a aceitação da vacinação infantil em toda a população dos EUA - Estudos publicados que mostram que vacinas causam autismo – Studies published showing vaccines cause autism
 Studies which show vaccines cause Autism: ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3878266/ ▪ http://ncbi.nlm.nih.gov/pubmed/21623535 ▪ http://ncbi.nlm.nih.gov/pubmed/25377033 ▪ http://ncbi.nlm.nih.gov/pubmed/24995277 ▪ http://ncbi.nlm.nih.gov/pubmed/12145534 ▪ http://ncbi.nlm.nih.gov/pubmed/21058170 ▪ http://ncbi.nlm.nih.gov/pubmed/22099159 ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3364648/ ▪ http://ncbi.nlm.nih.gov/pubmed/17454560 ▪ http://ncbi.nlm.nih.gov/pubmed/19106436 ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3774468/ ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3697751/ ▪ http://ncbi.nlm.nih.gov/pubmed/21299355 ▪ http://ncbi.nlm.nih.gov/pubmed/21907498 ▪ http://ncbi.nlm.nih.gov/pubmed/11339848 ▪ http://ncbi.nlm.nih.gov/pubmed/17674242 ▪ http://ncbi.nlm.nih.gov/pubmed/21993250 ▪ http://ncbi.nlm.nih.gov/pubmed/15780490 ▪ http://ncbi.nlm.nih.gov/pubmed/12933322 ▪ http://ncbi.nlm.nih.gov/pubmed/16870260 ▪ http://ncbi.nlm.nih.gov/pubmed/19043938 ▪ http://ncbi.nlm.nih.gov/pubmed/12142947 ▪ http://ncbi.nlm.nih.gov/pubmed/24675092 Relação causal entre… Leia mais: Estudos publicados que mostram que vacinas causam autismo – Studies published showing vaccines cause autism
Studies which show vaccines cause Autism: ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3878266/ ▪ http://ncbi.nlm.nih.gov/pubmed/21623535 ▪ http://ncbi.nlm.nih.gov/pubmed/25377033 ▪ http://ncbi.nlm.nih.gov/pubmed/24995277 ▪ http://ncbi.nlm.nih.gov/pubmed/12145534 ▪ http://ncbi.nlm.nih.gov/pubmed/21058170 ▪ http://ncbi.nlm.nih.gov/pubmed/22099159 ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3364648/ ▪ http://ncbi.nlm.nih.gov/pubmed/17454560 ▪ http://ncbi.nlm.nih.gov/pubmed/19106436 ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3774468/ ▪ http://ncbi.nlm.nih.gov/pmc/articles/PMC3697751/ ▪ http://ncbi.nlm.nih.gov/pubmed/21299355 ▪ http://ncbi.nlm.nih.gov/pubmed/21907498 ▪ http://ncbi.nlm.nih.gov/pubmed/11339848 ▪ http://ncbi.nlm.nih.gov/pubmed/17674242 ▪ http://ncbi.nlm.nih.gov/pubmed/21993250 ▪ http://ncbi.nlm.nih.gov/pubmed/15780490 ▪ http://ncbi.nlm.nih.gov/pubmed/12933322 ▪ http://ncbi.nlm.nih.gov/pubmed/16870260 ▪ http://ncbi.nlm.nih.gov/pubmed/19043938 ▪ http://ncbi.nlm.nih.gov/pubmed/12142947 ▪ http://ncbi.nlm.nih.gov/pubmed/24675092 Relação causal entre… Leia mais: Estudos publicados que mostram que vacinas causam autismo – Studies published showing vaccines cause autism - First editorial, translated into Russian. Хронический воспалительный процесс в мозге, – причина Расстройств Аутистического Спектра, требующая обязательного лечения
 Russian translator Dr Ph.D. Sementsov Vadim AlexandrovichInsta: @doctorsementsov Первое редакционное сообщение о причинах аутизма. Профессор Цицеро Гали Коимбра 2024 г. Аутизм был впервые описан Лео Каннером (детским психиатром из Медицинской школы Джона Хопкинса) в 1943 году, который назвал его «аутистическими расстройствами». Девять лет спустя стала очевидной степень тяжести этого поведенческого состояния, когда Американская психиатрическая ассоциация… Leia mais: First editorial, translated into Russian. Хронический воспалительный процесс в мозге, – причина Расстройств Аутистического Спектра, требующая обязательного лечения
Russian translator Dr Ph.D. Sementsov Vadim AlexandrovichInsta: @doctorsementsov Первое редакционное сообщение о причинах аутизма. Профессор Цицеро Гали Коимбра 2024 г. Аутизм был впервые описан Лео Каннером (детским психиатром из Медицинской школы Джона Хопкинса) в 1943 году, который назвал его «аутистическими расстройствами». Девять лет спустя стала очевидной степень тяжести этого поведенческого состояния, когда Американская психиатрическая ассоциация… Leia mais: First editorial, translated into Russian. Хронический воспалительный процесс в мозге, – причина Расстройств Аутистического Спектра, требующая обязательного лечения - Pagamentos da indústria farmacêutica e dispositivos médicos para revisores de pares dos EUA de grandes revistas médicas supera um bilhão de dólares
 Entre 2020 e 2022, a indústria farmacêutica destinou cerca de US$ 1,06 bilhão a revisores de grandes revistas médicas, incluindo The BMJ, JAMA, The Lancet e The New England Journal of Medicine. Essa informação já esteve postada por outra fonte do meio científico neste site, em 2020. Continua sendo denunciada em 2024. Fonte: JAMA. 2024;332(17):1480-1482.… Leia mais: Pagamentos da indústria farmacêutica e dispositivos médicos para revisores de pares dos EUA de grandes revistas médicas supera um bilhão de dólares
Entre 2020 e 2022, a indústria farmacêutica destinou cerca de US$ 1,06 bilhão a revisores de grandes revistas médicas, incluindo The BMJ, JAMA, The Lancet e The New England Journal of Medicine. Essa informação já esteve postada por outra fonte do meio científico neste site, em 2020. Continua sendo denunciada em 2024. Fonte: JAMA. 2024;332(17):1480-1482.… Leia mais: Pagamentos da indústria farmacêutica e dispositivos médicos para revisores de pares dos EUA de grandes revistas médicas supera um bilhão de dólares - ऑटिस्टिक मस्तिष्क एक अनुपचारित दीर्घकालिक सूजन प्रक्रिया से प्रभावित रहता है। इस वेबसाइट पर ऑटिज़्म के कारणों के बारे में यह पहला संपादकीय लेख है। – First editorial, hindi version: The autistic brain is affected by an untreated chronic inflammatory process
 Translated from English into Hindi by Dr. Renu Mahtani(अंग्रेजी से हिंदी में डॉ. रेणु महतानी द्वारा अनुवादित)Param Health Centre, Pune, Indiaparamhealth@gmail.com “ऑटिस्टिक मस्तिष्क एक अनुपचारित दीर्घकालिक सूजन प्रक्रिया से प्रभावित रहता है। इस वेबसाइट पर ऑटिज़्म के कारणों के बारे में यह पहला संपादकीय लेख है।” ऑटिज़्म का पहली बार वर्णन १९४३ में लियो कैनर… Leia mais: ऑटिस्टिक मस्तिष्क एक अनुपचारित दीर्घकालिक सूजन प्रक्रिया से प्रभावित रहता है। इस वेबसाइट पर ऑटिज़्म के कारणों के बारे में यह पहला संपादकीय लेख है। – First editorial, hindi version: The autistic brain is affected by an untreated chronic inflammatory process
Translated from English into Hindi by Dr. Renu Mahtani(अंग्रेजी से हिंदी में डॉ. रेणु महतानी द्वारा अनुवादित)Param Health Centre, Pune, Indiaparamhealth@gmail.com “ऑटिस्टिक मस्तिष्क एक अनुपचारित दीर्घकालिक सूजन प्रक्रिया से प्रभावित रहता है। इस वेबसाइट पर ऑटिज़्म के कारणों के बारे में यह पहला संपादकीय लेख है।” ऑटिज़्म का पहली बार वर्णन १९४३ में लियो कैनर… Leia mais: ऑटिस्टिक मस्तिष्क एक अनुपचारित दीर्घकालिक सूजन प्रक्रिया से प्रभावित रहता है। इस वेबसाइट पर ऑटिज़्म के कारणों के बारे में यह पहला संपादकीय लेख है। – First editorial, hindi version: The autistic brain is affected by an untreated chronic inflammatory process - Nos últimos anos, as indústrias de vacinas pagaram 35 bilhões de dólares em multas por falsificação de informações aos médicos, mas estão protegidas das consequências criminais
 *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens:
*Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens: - Análise de Robert Kennedy Jr. sobre a saúde nos Estados Unidos, especialmente de crianças e jovens, e as Agências Reguladoras – com legendas
 Leia as postagens pelo índice mensal: Leia as últimas postagens:
Leia as postagens pelo índice mensal: Leia as últimas postagens: - El cerebro autista se ve afectado por un proceso inflamatorio crónico no tratado. Primer Editorial en este Sitio sobre las causas del autismo, actualizado el 06/10/2024Traducido al español por Carmen Sanabria Ayala El autismo fue descrito por primera vez por Leo Kanner (psiquiatra infantil de la Escuela de Medicina Johns Hopkins) en 1943, quien lo llamó de “trastornos autistas” (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nueve años más tarde, la gravedad que esta condición conductual podría alcanzar se hizo evidente cuando la Asociación Americana de… Leia mais: El cerebro autista se ve afectado por un proceso inflamatorio crónico no tratado. Primer Editorial en este Sitio sobre las causas del autismo, actualizado el 06/10/2024
- Trabalhadores de trânsito dos EUA demitidos por recusarem vacina contra a COVID-19 receberão mais de US$ 1 milhão cada
 *Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens:
*Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens: - Romanian version: Procesul inflamator cronic, care dăunează creierului copiilor cu autism are o componentă autoimună incontestabilă. Al doilea Editorial de pe acest site despre cauzele autismuluiRomanian translation by Dr. Dominique Heidenfelder-Ambrosio Bazându-se pe logica luminată de publicațiile științifice, care s-au acumulat de-a lungul deceniilor, această serie de Editoriale își propune să ofere cititorului o cale de identificare a etiologiei (cauzei) autismului — o afecțiune al cărei cost anual, inclusiv cheltuielile directe și indirecte, ar putea depăși 460 de miliarde de… Leia mais: Romanian version: Procesul inflamator cronic, care dăunează creierului copiilor cu autism are o componentă autoimună incontestabilă. Al doilea Editorial de pe acest site despre cauzele autismului
- Autistični mozak zahvaćen je neliječenim kroničnim upalnim procesom. Prvi post na ovoj web stranici govori o uzrocima autizma
 Prijevod s engleskog na hrvatski Goranka Tomić i dr. Fedor Mataić Autizam je prvi opisao Leo Kanner (dječji psihijatar na Medicinskom fakultetu The Johns Hopkins) 1943. godine, koji ga je nazvao “autistični poremećaji”. (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Devet godina kasnije, ozbiljnost koju bi to stanje ponašanja moglo doseći postala je očigledna kada je Američka psihijatrijska udruga (APA) opisala… Leia mais: Autistični mozak zahvaćen je neliječenim kroničnim upalnim procesom. Prvi post na ovoj web stranici govori o uzrocima autizma
Prijevod s engleskog na hrvatski Goranka Tomić i dr. Fedor Mataić Autizam je prvi opisao Leo Kanner (dječji psihijatar na Medicinskom fakultetu The Johns Hopkins) 1943. godine, koji ga je nazvao “autistični poremećaji”. (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Devet godina kasnije, ozbiljnost koju bi to stanje ponašanja moglo doseći postala je očigledna kada je Američka psihijatrijska udruga (APA) opisala… Leia mais: Autistični mozak zahvaćen je neliječenim kroničnim upalnim procesom. Prvi post na ovoj web stranici govori o uzrocima autizma - Autismus – ein unbehandelter chronischer Entzündungsprozess des Gehirns. Erster Leitartikel auf dieser Website über die Ursachen von AutismusÜbersetzung aus der englischen Version: Prof. Dr. Nelson AnnunciatoNeurowissenschaftler Bewertet von:Dr. Melanie WeissWilhelmstraße 17342489 Wülfrathinfo@neurologie-wuelfrath.de Autismus wurde erstmals 1943 von Leo Kanner (einem Kinderpsychiater an der Johns Hopkins School of Medicine) beschrieben, der es als „autistische Störung “bezeichnete (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Neun Jahre später wurde die Schwere, die dieser Verhaltenszustand erreichen kann, deutlich, als die American Psychiatric… Leia mais: Autismus – ein unbehandelter chronischer Entzündungsprozess des Gehirns. Erster Leitartikel auf dieser Website über die Ursachen von Autismus
- The chronic inflammatory process that is damaging the brains of autistic children has an undeniable autoimmune component. The second editorial on this site on the causes of autism
 (Link to the first editorial: https://wp.me/pbW3AH-1DO -Link to the third editorial of this series: https://wp.me/pbW3AH-1Tx) Based on the logic enlightened by scientific publications that have accumulated over decades, this series of editorials aims to offer the reader a path to identifying the etiology (cause) of autism—a condition whose annual cost, including direct and indirect expenses,… Leia mais: The chronic inflammatory process that is damaging the brains of autistic children has an undeniable autoimmune component. The second editorial on this site on the causes of autism
(Link to the first editorial: https://wp.me/pbW3AH-1DO -Link to the third editorial of this series: https://wp.me/pbW3AH-1Tx) Based on the logic enlightened by scientific publications that have accumulated over decades, this series of editorials aims to offer the reader a path to identifying the etiology (cause) of autism—a condition whose annual cost, including direct and indirect expenses,… Leia mais: The chronic inflammatory process that is damaging the brains of autistic children has an undeniable autoimmune component. The second editorial on this site on the causes of autism - French version. Le cerveau des personnes autistes est affecté par un processus inflammatoire chronique non traité. Premier éditorial sur ce site web concernant les causes de l’autisme
 Traduction en français par Dr. Djamel Deramchi L’autisme a été décrit pour la première fois par Leo Kanner (pédopsychiatre à la faculté de médecine Johns Hopkins) en 1943, qui l’a qualifié de « troubles autistiques » (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Neuf ans plus tard, la gravité que cette condition comportementale pouvait atteindre est devenue évidente lorsque l’American Psychiatric Association (APA)… Leia mais: French version. Le cerveau des personnes autistes est affecté par un processus inflammatoire chronique non traité. Premier éditorial sur ce site web concernant les causes de l’autisme
Traduction en français par Dr. Djamel Deramchi L’autisme a été décrit pour la première fois par Leo Kanner (pédopsychiatre à la faculté de médecine Johns Hopkins) en 1943, qui l’a qualifié de « troubles autistiques » (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Neuf ans plus tard, la gravité que cette condition comportementale pouvait atteindre est devenue évidente lorsque l’American Psychiatric Association (APA)… Leia mais: French version. Le cerveau des personnes autistes est affecté par un processus inflammatoire chronique non traité. Premier éditorial sur ce site web concernant les causes de l’autisme - Il cervello del paziente autistico è affetto da un processo infiammatorio cronico non trattato. Su questo sito web primo Editoriale sulle cause dell’autismoTtradotto da Dottssa Giusi Manuele Specialista Endocrinochirurgia Protocollo Coimbra Italia L’autismo fu descritto per la prima volta da Leo Kanner (psichiatra infantile della Johns Hopkins School of Medicine) nel 1943, che lo chiamò “disturbo autistico” (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nove anni dopo, si comprese la gravità che questa condizione comportamentale poteva raggiungere e divenne evidente quando l’American Psychiatric… Leia mais: Il cervello del paziente autistico è affetto da un processo infiammatorio cronico non trattato. Su questo sito web primo Editoriale sulle cause dell’autismo
- Enquanto o Reino Unido debate a legalização do suicídio assistido, a prática aumenta drasticamente no Canadá
 Depois que as eleições de 4 de julho no Reino Unido fizeram de Keir Starmer o novo primeiro-ministro, o suicídio assistido virou notícia. A reportagem é de Charles Collins, publicada por Crux, 13-08-2024. No início deste ano, Starmer disse que estava pessoalmente comprometido em mudar as leis do país que tornam o suicídio assistido e a eutanásia ilegais. Os… Leia mais: Enquanto o Reino Unido debate a legalização do suicídio assistido, a prática aumenta drasticamente no Canadá
Depois que as eleições de 4 de julho no Reino Unido fizeram de Keir Starmer o novo primeiro-ministro, o suicídio assistido virou notícia. A reportagem é de Charles Collins, publicada por Crux, 13-08-2024. No início deste ano, Starmer disse que estava pessoalmente comprometido em mudar as leis do país que tornam o suicídio assistido e a eutanásia ilegais. Os… Leia mais: Enquanto o Reino Unido debate a legalização do suicídio assistido, a prática aumenta drasticamente no Canadá - Geração Z e Mortes do Desespero
 O professor de SPA analisa as tendências alarmantes do suicídio nacional, incluindo um aumento sem precedentes entre os jovens, e analisa possíveis determinantes. 10 de Agosto, 2023 Depois de cair por décadas, a taxa de suicídio nos Estados Unidos aumentou quase 40% desde 2000. Em “The Re-Emerging Suicide Crisis in the U.S.: Patterns, Causes and… Leia mais: Geração Z e Mortes do Desespero
O professor de SPA analisa as tendências alarmantes do suicídio nacional, incluindo um aumento sem precedentes entre os jovens, e analisa possíveis determinantes. 10 de Agosto, 2023 Depois de cair por décadas, a taxa de suicídio nos Estados Unidos aumentou quase 40% desde 2000. Em “The Re-Emerging Suicide Crisis in the U.S.: Patterns, Causes and… Leia mais: Geração Z e Mortes do Desespero - Estudo mostra taxas de suicídio e autolesão entre jovens em alta no Brasil
 Fonte: https://portal.fiocruz.br/en/news/2024/02/study-shows-suicide-and-self-injury-rates-rise-brazil 23/02/2024 Por Mariana Sebastião (Cidacs/Fiocruz Bahia) A taxa de suicídio entre os jovens cresceu 6% ao ano no Brasil entre 2011 e 2022. Por sua vez, as taxas de notificação para autolesões na faixa etária de 10 a 24 anos aumentaram 29% no mesmo período. O número foi maior em comparação com a… Leia mais: Estudo mostra taxas de suicídio e autolesão entre jovens em alta no Brasil
Fonte: https://portal.fiocruz.br/en/news/2024/02/study-shows-suicide-and-self-injury-rates-rise-brazil 23/02/2024 Por Mariana Sebastião (Cidacs/Fiocruz Bahia) A taxa de suicídio entre os jovens cresceu 6% ao ano no Brasil entre 2011 e 2022. Por sua vez, as taxas de notificação para autolesões na faixa etária de 10 a 24 anos aumentaram 29% no mesmo período. O número foi maior em comparação com a… Leia mais: Estudo mostra taxas de suicídio e autolesão entre jovens em alta no Brasil - Romanian version: Creierul autist este afectat de un proces inflamator cronic netratat. Primul editorial de pe acest site despre cauzele autismului
 Romanian translation by Dr. Dominique Heidenfelder-Ambrosio (English version: https://wp.me/pbW3AH-1DO) Autismul a fost descris pentru prima oară de către Leo Kanner (medic specialist în psihiatrie pediatrică de la Universitatea de Medicină John Hopkins) în 1943, care l-a denumit ca fiind „tulburări autiste”(https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nouă ani mai târziu, gravitatea pe care o poate atinge această condiție comportamentală, a… Leia mais: Romanian version: Creierul autist este afectat de un proces inflamator cronic netratat. Primul editorial de pe acest site despre cauzele autismului
Romanian translation by Dr. Dominique Heidenfelder-Ambrosio (English version: https://wp.me/pbW3AH-1DO) Autismul a fost descris pentru prima oară de către Leo Kanner (medic specialist în psihiatrie pediatrică de la Universitatea de Medicină John Hopkins) în 1943, care l-a denumit ca fiind „tulburări autiste”(https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nouă ani mai târziu, gravitatea pe care o poate atinge această condiție comportamentală, a… Leia mais: Romanian version: Creierul autist este afectat de un proces inflamator cronic netratat. Primul editorial de pe acest site despre cauzele autismului - Robert F. Kennedy Jr. compara as doenças infantis atuais com as da década de 80 – vídeo com legendas
 *Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens: *Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com
*Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com Leia as postagens pelo índice mensal: Leia as últimas postagens: *Todas as publicações deste site são escolha de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com - Estudo com 1,7 milhão de crianças e adolescentes que receberam a vacina Pfizer COVID encontrou miopericardite apenas em grupos vacinados -03.10.2024
 por Brenda Baletti, Ph.D.3 de outubro de 2024 “A vacina Pfizer-BioNTech COVID-19 forneceu a crianças e adolescentes na Inglaterra apenas cerca de 14 a 15 semanas de proteção contra testes positivos para o vírus, de acordo com um estudo de pré-impressão de mais de 1,7 milhão de crianças de 5 a 15 anos no Sistema… Leia mais: Estudo com 1,7 milhão de crianças e adolescentes que receberam a vacina Pfizer COVID encontrou miopericardite apenas em grupos vacinados -03.10.2024
por Brenda Baletti, Ph.D.3 de outubro de 2024 “A vacina Pfizer-BioNTech COVID-19 forneceu a crianças e adolescentes na Inglaterra apenas cerca de 14 a 15 semanas de proteção contra testes positivos para o vírus, de acordo com um estudo de pré-impressão de mais de 1,7 milhão de crianças de 5 a 15 anos no Sistema… Leia mais: Estudo com 1,7 milhão de crianças e adolescentes que receberam a vacina Pfizer COVID encontrou miopericardite apenas em grupos vacinados -03.10.2024 - The autistic brain is affected by an untreated chronic inflammatory process. First Editorial on this website about the causes of autism
 (Link to the third editorial of this series: https://wp.me/pbW3AH-1Tx) Autism was first described by Leo Kanner (a child psychiatrist at The Johns Hopkins School of Medicine) in 1943, who called it “autistic disorders” (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nine years later, the severity that this behavioral condition could reach became evident when the American Psychiatric Association (APA) described the… Leia mais: The autistic brain is affected by an untreated chronic inflammatory process. First Editorial on this website about the causes of autism
(Link to the third editorial of this series: https://wp.me/pbW3AH-1Tx) Autism was first described by Leo Kanner (a child psychiatrist at The Johns Hopkins School of Medicine) in 1943, who called it “autistic disorders” (https://www.autismtruths.org/pdf/Autistic%20Disturbances%20of%20Affective%20Contact%20-%20Leo%20Kanner.pdf). Nine years later, the severity that this behavioral condition could reach became evident when the American Psychiatric Association (APA) described the… Leia mais: The autistic brain is affected by an untreated chronic inflammatory process. First Editorial on this website about the causes of autism - Índice completo deste Site organizado por mês e ano – Complete index of this Site organized by month and year
 Ao acessar cada mês deste índice aqui publicado, será visto o título das postagens mensais com o acesso às mesmas. A partir do mês de julho de 2024, concentram-se as principais publicações sobre autismo. *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com… Leia mais: Índice completo deste Site organizado por mês e ano – Complete index of this Site organized by month and year
Ao acessar cada mês deste índice aqui publicado, será visto o título das postagens mensais com o acesso às mesmas. A partir do mês de julho de 2024, concentram-se as principais publicações sobre autismo. *Todas as publicações deste site são de exclusiva responsabilidade de seu criador e único administrador, Celso Galli Coimbra, OABRS 11352, e-mail cgcoimbra@gmail.com… Leia mais: Índice completo deste Site organizado por mês e ano – Complete index of this Site organized by month and year - Em junho de 2023, um dos principais governantes canadense pede desculpas aos não vacinados, “estávamos errados…” ela faz uma promessa sem precedentes …
 17 de junho de 2023 Danielle Smith, a atual primeira-ministra de Alberta, no Canadá, fez algo notável. Ela deu o passo ousado e sem precedentes de se desculpar com os canadenses não vacinados que enfrentaram tratamento injusto do governo durante a “pandemia”. Mas a Sra. Smith realmente foi além de apenas emitir um pedido de… Leia mais: Em junho de 2023, um dos principais governantes canadense pede desculpas aos não vacinados, “estávamos errados…” ela faz uma promessa sem precedentes …
17 de junho de 2023 Danielle Smith, a atual primeira-ministra de Alberta, no Canadá, fez algo notável. Ela deu o passo ousado e sem precedentes de se desculpar com os canadenses não vacinados que enfrentaram tratamento injusto do governo durante a “pandemia”. Mas a Sra. Smith realmente foi além de apenas emitir um pedido de… Leia mais: Em junho de 2023, um dos principais governantes canadense pede desculpas aos não vacinados, “estávamos errados…” ela faz uma promessa sem precedentes … - Vacinação obrigatória e confinamento incluídos no projeto de lei de saúde draconiano para Irlanda do Norte – Consulta pública em 14 de outubro de 2024
 Pela Dra. Elizabeth Evans, em 26 de setembro de 2024 CEO da Medical Freedom Alliance do Reino Unido O governo da Irlanda do Norte está atualmente realizando uma consulta pública sobre o Projeto de Lei de Saúde Pública Orwelliano. O novo projeto de lei busca atualizar e fortalecer a Lei de Saúde Pública existente e… Leia mais: Vacinação obrigatória e confinamento incluídos no projeto de lei de saúde draconiano para Irlanda do Norte – Consulta pública em 14 de outubro de 2024
Pela Dra. Elizabeth Evans, em 26 de setembro de 2024 CEO da Medical Freedom Alliance do Reino Unido O governo da Irlanda do Norte está atualmente realizando uma consulta pública sobre o Projeto de Lei de Saúde Pública Orwelliano. O novo projeto de lei busca atualizar e fortalecer a Lei de Saúde Pública existente e… Leia mais: Vacinação obrigatória e confinamento incluídos no projeto de lei de saúde draconiano para Irlanda do Norte – Consulta pública em 14 de outubro de 2024 - Em 29.08.2024, a Food and Drug Administration (FDA) aprovou a vacina contra a varíola do macaco, mesmo reconhecendo que é perigosa
 O vírus da vacina aprovada nos EUA pode atacar as pessoas próximas à pessoa que se vacinou. “Mesmo o guia de medicamentos do regulador para a injeção admite que pode causar mais dano do que benefício.” ‘Para piorar as coisas, o ACAM2000 “pode causar complicações graves em indivíduos vacinados e em seus contatos próximos aos… Leia mais: Em 29.08.2024, a Food and Drug Administration (FDA) aprovou a vacina contra a varíola do macaco, mesmo reconhecendo que é perigosa
O vírus da vacina aprovada nos EUA pode atacar as pessoas próximas à pessoa que se vacinou. “Mesmo o guia de medicamentos do regulador para a injeção admite que pode causar mais dano do que benefício.” ‘Para piorar as coisas, o ACAM2000 “pode causar complicações graves em indivíduos vacinados e em seus contatos próximos aos… Leia mais: Em 29.08.2024, a Food and Drug Administration (FDA) aprovou a vacina contra a varíola do macaco, mesmo reconhecendo que é perigosa - O processo inflamatório crônico que está lesando o cérebro das crianças autistas possui um inegável componente autoimune – Segundo Editorial deste Site sobre as causas do autismo
 (Link do primeiro editorial: https://wp.me/pbW3AH-1yv) Fundamentando-se na lógica iluminada por publicações científicas que vêm se acumulando ao longo de décadas, esta sequência de Editoriais almeja oferecer ao leitor um trajeto para a identificação da etiologia (causa) do autismo – uma doença cujo custo anual entre despesas diretas e indiretas pode atingir mais de 460 bilhões… Leia mais: O processo inflamatório crônico que está lesando o cérebro das crianças autistas possui um inegável componente autoimune – Segundo Editorial deste Site sobre as causas do autismo
(Link do primeiro editorial: https://wp.me/pbW3AH-1yv) Fundamentando-se na lógica iluminada por publicações científicas que vêm se acumulando ao longo de décadas, esta sequência de Editoriais almeja oferecer ao leitor um trajeto para a identificação da etiologia (causa) do autismo – uma doença cujo custo anual entre despesas diretas e indiretas pode atingir mais de 460 bilhões… Leia mais: O processo inflamatório crônico que está lesando o cérebro das crianças autistas possui um inegável componente autoimune – Segundo Editorial deste Site sobre as causas do autismo - Vacina de mRNA COVID-19 ligada a cicatrizes miocárdicas em adolescentes e adultos jovens – 11.09.2024 eClinicalMedicine
 “Novo estudo revela maior prevalência de cicatrizes cardíacas em homens jovens após a vacinação de mRNA contra COVID-19, levantando preocupações sobre os resultados a longo prazo, apesar dos sintomas iniciais leves de miocardite associada à vacina” “Embora as implicações e o prognóstico geral para lesão miocárdica associada ao C-VAM em pacientes jovens não sejam claros,… Leia mais: Vacina de mRNA COVID-19 ligada a cicatrizes miocárdicas em adolescentes e adultos jovens – 11.09.2024 eClinicalMedicine
“Novo estudo revela maior prevalência de cicatrizes cardíacas em homens jovens após a vacinação de mRNA contra COVID-19, levantando preocupações sobre os resultados a longo prazo, apesar dos sintomas iniciais leves de miocardite associada à vacina” “Embora as implicações e o prognóstico geral para lesão miocárdica associada ao C-VAM em pacientes jovens não sejam claros,… Leia mais: Vacina de mRNA COVID-19 ligada a cicatrizes miocárdicas em adolescentes e adultos jovens – 11.09.2024 eClinicalMedicine - Flórida desaconselha o uso da vacina de mRNA COVID no outono e inverno
 “A declaração levantou preocupações adicionais sobre a decisão do governo federal de não exigir que os fabricantes demonstrem a eficácia dos reforços na prevenção de hospitalizações ou morte.“ “A Flórida também alertou sobre riscos de segurança, incluindo o potencial de circulação prolongada de mRNA e proteína spike, risco potencial aumentado de infecções respiratórias, doenças autoimunes… Leia mais: Flórida desaconselha o uso da vacina de mRNA COVID no outono e inverno
“A declaração levantou preocupações adicionais sobre a decisão do governo federal de não exigir que os fabricantes demonstrem a eficácia dos reforços na prevenção de hospitalizações ou morte.“ “A Flórida também alertou sobre riscos de segurança, incluindo o potencial de circulação prolongada de mRNA e proteína spike, risco potencial aumentado de infecções respiratórias, doenças autoimunes… Leia mais: Flórida desaconselha o uso da vacina de mRNA COVID no outono e inverno - A criança como sujeito de experimentação científica: uma análise histórica dos aspectos éticos
 Algumas histórias exemplares da crueldade em nome da ciência, dados extraídos da tese de doutoramento de Joaquim Antônio César Mota: “A criança como sujeito de experimentação científica: uma análise histórica dos aspectos éticos”, UFMG, agosto de 1998. O processo de diagnosticar e tratar as moléstias sempre teve aspectos experimentais. Começou com a aplicação deste ou daquele… Leia mais: A criança como sujeito de experimentação científica: uma análise histórica dos aspectos éticos
Algumas histórias exemplares da crueldade em nome da ciência, dados extraídos da tese de doutoramento de Joaquim Antônio César Mota: “A criança como sujeito de experimentação científica: uma análise histórica dos aspectos éticos”, UFMG, agosto de 1998. O processo de diagnosticar e tratar as moléstias sempre teve aspectos experimentais. Começou com a aplicação deste ou daquele… Leia mais: A criança como sujeito de experimentação científica: uma análise histórica dos aspectos éticos - El cerebro del autista está afectado por un proceso inflamatorio crónico no tratado. Primer Editorial de este Site sobre las causas del autismoUNA TRAGEDIA ENCUBIERTA* (No se permite la reproducción de este texto, publicado aquí en https://protocolocoimbradrcicerogalli.com, el 13 de agosto de 2024, en ninguna de las redes sociales y en cualquier otro medio. Publicación en portugués: https://wp.me/pbW3AH-1yv ) [Se hace necesario la dosificación de los niveles circulantes de la enzima enolasa neuronal específica.] El autismo fue… Leia mais: El cerebro del autista está afectado por un proceso inflamatorio crónico no tratado. Primer Editorial de este Site sobre las causas del autismo
- Em 2018, o “Padrinho das Vacinas” no mundo, Dr. Stanley Plotkin, declara ter testado vacinas experimentais em órfãos, em deficientes mentais, em bebês de mães presas e em população de domínios coloniais, alcançando 1.000.000 de pessoas – vídeo com legendas
 Polêmica: |Em carta ao New England Journal Medicine, o Dr. Plotkin expressou preocupações sobre a realização de experimentos em adultos e crianças “plenamente funcionais” que são potenciais contribuintes para a sociedade, em vez de usar “sujeitos humanos que são humanos na forma, mas não no potencial social” (A., Plotkin, Stanley; P., Moore, Jerry, “Ethics of… Leia mais: Em 2018, o “Padrinho das Vacinas” no mundo, Dr. Stanley Plotkin, declara ter testado vacinas experimentais em órfãos, em deficientes mentais, em bebês de mães presas e em população de domínios coloniais, alcançando 1.000.000 de pessoas – vídeo com legendas
Polêmica: |Em carta ao New England Journal Medicine, o Dr. Plotkin expressou preocupações sobre a realização de experimentos em adultos e crianças “plenamente funcionais” que são potenciais contribuintes para a sociedade, em vez de usar “sujeitos humanos que são humanos na forma, mas não no potencial social” (A., Plotkin, Stanley; P., Moore, Jerry, “Ethics of… Leia mais: Em 2018, o “Padrinho das Vacinas” no mundo, Dr. Stanley Plotkin, declara ter testado vacinas experimentais em órfãos, em deficientes mentais, em bebês de mães presas e em população de domínios coloniais, alcançando 1.000.000 de pessoas – vídeo com legendas - Pesquisador denuncia problemas de integridade de dados em teste de vacina Covid-19 da Pfizer
 “Um diretor regional que era empregado na organização de pesquisa Ventavia Research Group disse ao The BMJ que a empresa falsificou dados, revelou pacientes, empregou vacinadores inadequadamente treinados e demorou a acompanhar os eventos adversos relatados no ensaio clínico de fase III da Pfizer.” “Revelações de práticas ruins em uma empresa de pesquisa contratada que… Leia mais: Pesquisador denuncia problemas de integridade de dados em teste de vacina Covid-19 da Pfizer
“Um diretor regional que era empregado na organização de pesquisa Ventavia Research Group disse ao The BMJ que a empresa falsificou dados, revelou pacientes, empregou vacinadores inadequadamente treinados e demorou a acompanhar os eventos adversos relatados no ensaio clínico de fase III da Pfizer.” “Revelações de práticas ruins em uma empresa de pesquisa contratada que… Leia mais: Pesquisador denuncia problemas de integridade de dados em teste de vacina Covid-19 da Pfizer - Associação de cientistas da América Latina firmou declaração fundamentada em 2020 contrária à vacinação Covid-19, especialmente a forçada
 “A legislação é feita para favorecer a indústria farmacêutica multinacional através de imunidade jurídica e exoneração de responsabilidade por possíveis danos. Aceitam acordos de confidencialidade que protegem dados técnicos básicos e recebem recursos públicos substanciais.” Declaração da UCCSNAL sobre novas vacinas genéticas ou transgênicas no contexto da SARS COVID19 3 de novembro de 2020 |… Leia mais: Associação de cientistas da América Latina firmou declaração fundamentada em 2020 contrária à vacinação Covid-19, especialmente a forçada
“A legislação é feita para favorecer a indústria farmacêutica multinacional através de imunidade jurídica e exoneração de responsabilidade por possíveis danos. Aceitam acordos de confidencialidade que protegem dados técnicos básicos e recebem recursos públicos substanciais.” Declaração da UCCSNAL sobre novas vacinas genéticas ou transgênicas no contexto da SARS COVID19 3 de novembro de 2020 |… Leia mais: Associação de cientistas da América Latina firmou declaração fundamentada em 2020 contrária à vacinação Covid-19, especialmente a forçada - Desenvolvimento de vacinas para covid-19 é um jogada histórica de lucro para a grande indústria farmacêutica
 Por Silvia Ribeiro Tradução: Luiza Mançano – Agência Latino-americana de Informação (ALAI) O desenvolvimento de vacinas para covid-19 é uma jogada histórica de lucro para a grande indústria farmacêutica. Não apenas por alcançarem eficácia e vendê-la, mas também devido à especulação financeira desde antes e, sobretudo, graças aos enormes subsídios governamentais que estão recebendo. Segundo… Leia mais: Desenvolvimento de vacinas para covid-19 é um jogada histórica de lucro para a grande indústria farmacêutica
Por Silvia Ribeiro Tradução: Luiza Mançano – Agência Latino-americana de Informação (ALAI) O desenvolvimento de vacinas para covid-19 é uma jogada histórica de lucro para a grande indústria farmacêutica. Não apenas por alcançarem eficácia e vendê-la, mas também devido à especulação financeira desde antes e, sobretudo, graças aos enormes subsídios governamentais que estão recebendo. Segundo… Leia mais: Desenvolvimento de vacinas para covid-19 é um jogada histórica de lucro para a grande indústria farmacêutica - Como a indústria farmacêutica compra médicos – How drug companies buy doctors
 As afirmações do Professor Dr. Peter Gøtzsche sobre este assunto crítico na Medicina não significam que ele está referindo-se a todos os médicos. Portanto, suas explicações referem-se a profissionais não éticos de qualquer nacionalidade, e profissionais não éticos podem ser encontrados em todas as profissões. O problema surge quando se confunde defender uma profissão com… Leia mais: Como a indústria farmacêutica compra médicos – How drug companies buy doctors
As afirmações do Professor Dr. Peter Gøtzsche sobre este assunto crítico na Medicina não significam que ele está referindo-se a todos os médicos. Portanto, suas explicações referem-se a profissionais não éticos de qualquer nacionalidade, e profissionais não éticos podem ser encontrados em todas as profissões. O problema surge quando se confunde defender uma profissão com… Leia mais: Como a indústria farmacêutica compra médicos – How drug companies buy doctors - “Oftalmologistas agora são eticamente obrigados a denunciar as vacinas COVID-19, já que 20.000 novos distúrbios oculares são relatados”
 “O holocausto da vacina covid está destruindo a audição e a visão das pessoas” 24 julho 2024 by Lance Johnson “Distúrbios oculares relacionados à vacina (do processador em andamento) relatados pela primeira vez há dois anos, em maio de 2021 “Publicado pela primeira vez em 6 de maio de 2021” “Em apenas alguns meses, a… Leia mais: “Oftalmologistas agora são eticamente obrigados a denunciar as vacinas COVID-19, já que 20.000 novos distúrbios oculares são relatados”
“O holocausto da vacina covid está destruindo a audição e a visão das pessoas” 24 julho 2024 by Lance Johnson “Distúrbios oculares relacionados à vacina (do processador em andamento) relatados pela primeira vez há dois anos, em maio de 2021 “Publicado pela primeira vez em 6 de maio de 2021” “Em apenas alguns meses, a… Leia mais: “Oftalmologistas agora são eticamente obrigados a denunciar as vacinas COVID-19, já que 20.000 novos distúrbios oculares são relatados” - “Desde 1986, com a isenção legal de responsabilidades por danos à saúde e mortes, a indústria das vacinas cresce em um ambiente burocrático imensamente falho e corrupto”
 Proibida a divulgação deste texto fora deste Site. Para referência indicar link. Em 12 de setembro de 2023, o advogado Aaron Siri presta depoimento como testemunha em audiência pública perante o Comitê Ad-Hoc de Assuntos Médicos do Senado da Carolina do Sul sobre o DHEC (Departamento de Saúde e Controle Ambiental da Carolina do Sul). Nessa… Leia mais: “Desde 1986, com a isenção legal de responsabilidades por danos à saúde e mortes, a indústria das vacinas cresce em um ambiente burocrático imensamente falho e corrupto”
Proibida a divulgação deste texto fora deste Site. Para referência indicar link. Em 12 de setembro de 2023, o advogado Aaron Siri presta depoimento como testemunha em audiência pública perante o Comitê Ad-Hoc de Assuntos Médicos do Senado da Carolina do Sul sobre o DHEC (Departamento de Saúde e Controle Ambiental da Carolina do Sul). Nessa… Leia mais: “Desde 1986, com a isenção legal de responsabilidades por danos à saúde e mortes, a indústria das vacinas cresce em um ambiente burocrático imensamente falho e corrupto”

